ņä£ ļĪĀ
ņŗ£ņŖżĒöīļØ╝Ēŗ┤(cisplatin)ņØĆ ņóģņ¢æņäĖĒżņØś ņ”ØņŗØņØä ņ¢ĄņĀ£ĒĢśļŖö ĒĢŁņĢöņĀ£ļĪ£ņä£ ļæÉĻ▓ĮļČĆ ņĢģņä▒ņóģņ¢æ ļ░Å ņŚ¼ļ¤¼ Ļ│ĀĒśĢņĢö(ļé£ņåīņĢö, Ļ│ĀĒÖśņĢö, ļ░®Ļ┤æņĢö, ĒÅÉņĢö ļō▒)ņØś ņ╣śļŻīņŚÉ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ļŗż[1,2]. ņØ┤ļŖö DNAņŚÉ ļ╣äĻ░ĆņŚŁņĀüņ£╝ļĪ£ ļ░śņØæĒĢśņŚ¼ ĻĄ¼ņĢäļŗīĻĖ░ņØś ļéśņäĀ Ļ░Ćļŗźļōż ņé¼ņØ┤ņŚÉņä£ņØś ņāüĒśĖ ņŚ░Ļ▓░ņØä ĒåĄĒĢ┤ ņäĖĒż ĒīīĻ┤┤ļź╝ ņ£Āļ░£ĒĢśļŖö ņĢĮļ¼╝ļĪ£ņä£ ĒĢŁņĢö ĒÜ©Ļ│╝ ņÖĖņŚÉ Ļ░Ćļ│ŹĻ▓īļŖö ĻĄ¼ņŚŁ, ĻĄ¼ĒåĀ ļō▒ņØś ļČĆņ×æņÜ® ņØ╝ņ£╝Ēéżļ®░, ņŗĀļÅģņä▒, ņŗĀĻ▓ĮļÅģņä▒ ļ░Å ņØ┤ļÅģņä▒ ļō▒ņØś ņśüĻĄ¼ņĀüņØĖ ļČĆņ×æņÜ® ļśÉĒĢ£ ņ£Āļ░£ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[1,2]. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ£ ļČĆņ×æņÜ®ņØĆ ĒŖ╣Ē׳ ņåīņĢäņŚÉņä£ ņŗ¼ĒĢśļ®░, Ēł¼ņŚ¼ ļ░øļŖö ņåīņĢäņØś 60% ņØ┤ņāüņØ┤ ņŗĀĻĖ░ļŖźņןņĢĀļź╝ Ļ▓ĮĒŚśĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░, ņĢĮ 60% ņØ┤ņāüņØś ņåīņĢäņŚÉņä£ ņśüĻĄ¼ņĀüņØĖ ņ▓ŁļĀźņĀĆĒĢśĻ░Ć ņ£Āļ░£ļÉ£ļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņŚłļŗż[3-5]. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ£ ļé£ņ▓ŁņØĆ ņ¢æņĖĪņä▒ņ£╝ļĪ£ ņ¦äĒ¢ēĒĢśļŖö ļ╣äĻ░ĆņŚŁņĀü Ļ░ÉĻ░üņŗĀĻ▓Įņä▒ ļé£ņ▓ŁņØś ĒśĢĒā£ļĪ£, ņĀä ņŚ░ļĀ╣ļīĆņŚÉņä£ņØś ņ£Āļ│æļźĀņØĆ ļ¼ĖĒŚīņŚÉ ļö░ļØ╝ ļŗżņ¢æĒĢśņŚ¼ 11%-97% ņĀĢļÅäņØ┤ļ®░, ĒÅēĻĘĀ 62%ņØś ļ░£ņāØļ╣łļÅäļź╝ ļ│┤ņØ┤ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[6,7]. ņØ┤ļŖö Ēł¼ņŚ¼ ļ░®ļ▓Ģ, Ēł¼ņŚ¼ ņÜ®ļ¤ē, ņ▓┤ļé┤ ļłäņĀü ņÜ®ļ¤ē, ĒÖśņ×ÉņØś ļéśņØ┤, ļŗżļźĖ ņØ┤ļÅģņä▒ ņĢĮļ¼╝ņØś ļ│æņÜ®Ēł¼ņŚ¼ ļ░Å ļæÉĻ▓ĮļČĆ ļ░®ņé¼ņäĀ ņ╣śļŻī ļō▒Ļ│╝ ļ░ĆņĀæĒĢ£ Ļ┤ĆļĀ©ņØ┤ ņ׳ņ£╝ļ®░, Ļ│ĀņØīņŚŁļīĆļČĆĒä░ ņ¦äĒ¢ēĒĢśļŖö ļé£ņ▓Ł ņÖĖņŚÉļÅä ņĢĮ 2%-36%ņŚÉņä£ ņØ┤ļ¬ģņØä ļÅÖļ░śĒĢśĻĖ░ļÅä ĒĢ£ļŗż[8]. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ĒśłĻ┤ĆņĪ░ņŚÉņä£ adenylate cyclase ņ×æņÜ®ņØä ņ¢ĄņĀ£ĒĢśņŚ¼, ņ£Āļ¬©ņäĖĒżņØś ņłśņÜ®ņ▓┤ ņĀäļźś(receptor current)ļź╝ Ļ░Éņåīņŗ£ņ╝£ ļé£ņ▓ŁņØä ņ£Āļ░£ĒĢ£ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░, ņÖĖņ£Āļ¬©ņäĖĒż ļ░Å ņ¦Ćņ¦ĆņäĖĒż ļō▒ņØś ĒśĢĒā£ĒĢÖņĀü ņØ┤ņāü ļśÉĒĢ£ ņ£Āļ░£ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņŚłļŗż[9]. ņØ┤ņ▓śļ¤╝ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØä ņÖĆņÜ░ņØś ņ¦üņĀæņĀü ņåÉņāüņ£╝ļĪ£ ņäżļ¬ģĒĢśļŖö Ļ▓āņØ┤ ņØ╝ļ░śņĀüņØ┤ļéś, ĒĢŁņĢöņĀ£ļĪ£ņä£ņØś ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś Ēæ£ņĀüņØĆ ņ”ØņŗØĒĢśļŖö ņóģņ¢æ ņäĖĒżņØś ĒĢĄ DNAņØĖļŹ░ ļ░śĒĢ┤, ņ”ØņŗØĒĢśņ¦Ć ņĢŖļŖö ņÖĆņÜ░ņäĖĒżĻ░Ć ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ņśüĒ¢źņØä ļ░øļŖö ĻĖ░ņĀäņØĆ ņĢäņ¦ü ļ¬ģĒÖĢĒ׳ ņØ┤ĒĢ┤ļÉśņ¦Ć ņĢŖļŖöļŗż. ļö░ļØ╝ņä£ Ēśäņ×¼Ļ╣īņ¦Ć ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØś ļ¬ģĒÖĢĒĢ£ ļ░£ņāØ ļČĆņ£äņÖĆ ĻĖ░ņĀäņŚÉ ļīĆĒĢ┤ņä£ļŖö ļģ╝ļ×ĆņØ┤ ņ׳ņ£╝ļ®░, ņØ┤ļź╝ ņśłļ░®ĒĢĀ ņłś ņ׳ļŖö FDA ņŖ╣ņØĖ ņĢĮļ¼╝ļÅä ņŚåļŖö ņāüĒā£ņØ┤ļŗż[9].
ļ│Ė ņóģņäżņŚÉņä£ļŖö Ēśäņ×¼Ļ╣īņ¦Ć ņĢīļĀżņ¦ä ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ĒĢŁņĢöņĀ£ņØś ņØ┤ļÅģņä▒ ĻĖ░ņĀäņØä ņĀĢļ”¼ĒĢśņŚ¼, ņØ┤ļÅģņä▒ņ£╝ļĪ£ļČĆĒä░ ņÖĆņÜ░ņ£Āļ¬©ņäĖĒżļź╝ ļ│┤ĒśĖĒĢśĻĖ░ ņ£äĒĢ£ ņŚ░ĻĄ¼ņØś ņ┤łņäØņ£╝ļĪ£ņä£ ļČäņ×É Ēæ£ņĀüņØś ņØ┤ĒĢ┤ļź╝ ļÅĢĻ│Āņ×É ĒĢ£ļŗż.
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ĒĢŁņĢöņ×æņÜ® ĻĖ░ņĀä
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ 1844ļģä ņØ┤Ēāłļ”¼ņĢä ĒÖöĒĢÖņ×ÉņØĖ Michele PeyroneņŚÉ ņØśĒĢ┤ ņ▓śņØī ĒĢ®ņä▒ļÉśņŚłņ£╝ļ®░, 1966ļģä ļ»Ėņŗ£Ļ▒┤ ļīĆĒĢÖņøÉņØś ņāØļ¼╝ĒĢÖņ×ÉņØĖ Barnett RosenbergĻ░Ć ņ¦äĒĢĄņäĖĒż ļČäņŚ┤ņŚÉ ļīĆĒĢ£ ņ×ÉĻĖ░ņןņØś ņśüĒ¢źņØä ņĪ░ņé¼ĒĢśļŹś ņżæ ņÜ░ņŚ░Ē׳ ņ×ÉĻĖ░ņןņŚÉņä£ ņ▓ĀņØś ļ░░ņŚ┤ņØ┤ ņäĖĒżļČäņŚ┤ņŚÉņä£ ļ│╝ ņłś ņ׳ļŖö ņ£Āņé¼ļČäņŚ┤ ļ░®ņČöņØś ļ░░ņŚ┤Ļ│╝ ņ£Āņé¼ĒĢśļŗżļŖö Ļ▓āņØä ļ░£Ļ▓¼ĒĢśļ®┤ņä£ ņØ┤ņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņŗ£ņ×æļÉśņŚłļŗż[10,11]. ĻĘĖļŖö ĒøäņåŹ ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ņÜ®ĒĢ┤ļÉ£ ļ░▒ĻĖłņØ┤ ņĢöļ¬©ļŖä ļ░Å ņŚ╝ĒÖöļ¼╝ņØ┤ņś©Ļ│╝ ļ░śņØæĒĢśņŚ¼ ņäĖĒż ļČäņŚ┤ņØä ņ¢ĄņĀ£ĒĢśļŖö ņŚŁĒĢĀņØä ĒĢ£ļŗżļŖö Ļ▓āņØä ļ░£Ļ▓¼ĒĢśņśĆļŖöļŹ░, ņØ┤ ĒÖöĒĢ®ļ¼╝ņØ┤ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØ┤ļŗż[12,13]. ņØ┤Ēøä ļ░▒Ēśłļ│æĻ│╝ ņ£Īņóģ(sarcoma) ļ¦łņÜ░ņŖżļ¬©ļŹĖ ņŗżĒŚśņØä ĒåĄĒĢ┤ ņØ┤ ĒÖöĒĢ®ļ¼╝ņØś ĒĢŁ ņóģņ¢æ ĒÖ£ņä▒ņØä ĒÖĢņØĖĒĢ©ņ£╝ļĪ£ņŹ©, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ Ēśäņ×¼ ļæÉĻ▓ĮļČĆņĢö, ņŗØļÅäņĢö, ĒÅÉņĢö, Ļ│ĀĒÖśņĢö, ļé£ņåīņĢö, ņ×ÉĻČüĻ▓ĮļČĆņĢö ļō▒ ļŗżņ¢æĒĢ£ Ļ│ĀĒśĢ ņóģņ¢æņØś ņ╣śļŻīņŚÉ ļäÉļ”¼ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ļŗż[13-15].
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ļŗżņ¢æĒĢ£ ĻĖ░ņĀäņ£╝ļĪ£ ņĢöņäĖĒżļź╝ ņĀ£Ļ▒░ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖöļŹ░, ņóģņ¢æņäĖĒż DNAņÖĆņØś Ļ▓░ĒĢ® ļ░Å ļČĆĻ░Ćļ¼╝ ĒśĢņä▒(adduct formation)ņØ┤ ņóģņ¢æ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ņŻ╝ņÜö Ēæ£ņĀüņØĖ Ļ▓āņ£╝ļĪ£ ņŚ¼Ļ▓©ņ¦äļŗż. ņŗ£ņŖżĒöīļØ╝Ēŗ┤Ļ│╝ ņóģņ¢æņäĖĒż DNAņØś Ļ▓░ĒĢ®ņØĆ ņĀäņé¼ņÖĆ DNA ļ│ĄņĀ£ļź╝ ņ¢ĄņĀ£ĒĢśĻ│Ā, ņŗ£ņŖżĒöīļØ╝Ēŗ┤-DNA ļČĆĻ░Ćļ¼╝ņØś ļ│ĄĻĄ¼Ļ░Ć ņĀ£ļīĆļĪ£ ņØ┤ļŻ©ņ¢┤ņ¦Ćņ¦Ć ņĢŖņ£╝ļ®┤, ņĢöņäĖĒżņØś ņäĖĒżņ×Éļ®Ėņé¼(apoptosis)Ļ░Ć ņØ╝ņ¢┤ļéśļ®┤ņä£ ņäĖĒżĻ░Ć ņé¼ļ®ĖĒĢ£ļŗż[16-18]. ļśÉĒĢ£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņóģņ¢æņäĖĒżņØś DNAņŚÉ Ļ▓░ĒĢ®ĒĢśņŚ¼ Ļ░ĢļĀźĒĢ£ ņé░ĒÖö ņŖżĒŖĖļĀłņŖż ļ░śņØæņØä ņāØņä▒ĒĢśļ®░, Ļ┤┤ņé¼(necoptosis) ļ░Å ferroptosisļź╝ ĒåĄĒĢ┤ ņČöĻ░ĆņĀüņØĖ ņäĖĒżļÅģņä▒ ĻĖ░ņĀäņ£╝ļĪ£ ņäĖĒżņé¼ļ®ĖņØä ņØ╝ņ£╝ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ļŗż[19-22].
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒
ņĀäņŗĀĒł¼ņŚ¼ļÉ£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņ┤łĻĖ░ņŚÉļŖö ņÖĆņÜ░ņØś Ļ│ĀņŻ╝ĒīīņśüņŚŁņŚÉņä£ ņÖĖņ£Āļ¬©ņäĖĒżņØś ņé¼ļ®ĖņØä ņØ╝ņ£╝Ēéżņ¦Ćļ¦ī, ņ¦äĒ¢ēļÉśļ®┤ ņĀĆņŻ╝ĒīīņśüņŚŁņ£╝ļĪ£ ņäĖĒż ņé¼ļ®ĖņØ┤ ņ¦äĒ¢ēļÉ£ļŗż[8,23,24]. ļö░ļØ╝ņä£ ņ¦ĆņåŹņĀüņØĖ ņØ┤ļÅģņåīņØś ņ▓┤ļé┤ ļłäņĀüļ¤ēņØś ņ”ØĻ░ĆļŖö ņĀäņŻ╝Ēīīņłś ņśüņŚŁņ£╝ļĪ£ņØś ņśüĻĄ¼ņĀüņØĖ ņ▓ŁļĀź ņåÉņŗżņØś ņ£äĒŚśņä▒ ņ”ØĻ░Ćļź╝ ņØśļ»ĖĒĢ£ļŗż[23,25]. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņŻ╝ļĪ£ ņ£Āļ¬©ņäĖĒżņŚÉ ņ¦üņĀæņĀüņØĖ ņåÉņāüņØä ļ»Ėņ╣śļŖö Ļ▓āņ£╝ļĪ£ ņל ņĢīļĀżņĀĖņ׳ņ£╝ļéś, ņ£Āļ¬©ņäĖĒż ņÖĖņŚÉļÅä ņ¦Ćņ¦ĆņäĖĒż, ļéśņäĀņŗĀĻ▓ĮņĀł ņäĖĒżņŚÉļÅä ņśüĒ¢źņØä ņŻ╝ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņŚłļŗż[26-28]. ņØ┤ļÅģņä▒ņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ņØś ļīĆļČĆļČäņØ┤ ņĀĢņāü ņÖĆņÜ░ļź╝ ņØ┤ņÜ®ĒĢ£ ņŚ░ĻĄ¼ļĪ£ ņ¦äĒ¢ēļÉśĻĖ░ ļĢīļ¼ĖņŚÉ, ņØ┤ļÅģņä▒ņØś ņØ╝ņ░© ļČĆņ£äņŚÉ ļīĆĒĢ£ Ļ▓░ņĀĢņØĆ ņēĮņ¦Ć ņĢŖļŗż. ņØ╝ļČĆ ņŚ░ĻĄ¼ņŚÉņä£ņØś ļ│ĄĒĢ®ĒÖ£ļÅÖņĀäņ£ä(compound action potential, CAP)ņÖĆ ņ▓Łņä▒ļćīĻ░äļ░śņØæ(auditory brainstem response, ABR) ņŚŁņ╣ś ņ”ØĻ░Ć Ļ▓░Ļ│╝ļŖö ļéśņäĀ ņŗĀĻ▓ĮņĀłņØś ņåÉņāüņØä ļ░śņśüĒĢśļŖöļŹ░, ņØ┤ļŖö ņÖĖņ£Āļ¬©ņäĖĒżņÖĆ ļÅÖņŗ£ņŚÉ ņĀ£ 1ĒśĢ ļéśņä▒ ņŗĀĻ▓ĮņĀłņØś ņłśņ┤ł ņåÉņŗżĻ│╝ Ļ┤ĆļĀ©ņØ┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż[29]. ļśÉĒĢ£ ABR ņŚŁņ╣ś ņ”ØĻ░Ć Ļ▓░Ļ│╝ļŖö ļé┤ļ”╝ĒöäņØś Ca2+ ļ░Å K+ ļåŹļÅäņØś Ļ░ÉņåīļĪ£ ņØĖĒĢ£ Ļ▓░Ļ│╝ņØ╝ ņłś ņ׳ļŖöļŹ░, ņØ┤ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ£ ņÖĆņÜ░ ņĖĪļ▓ĮņØś Na+/K+-ATPase ĒÖ£ņä▒ Ļ░ÉņåīņÖĆ Ļ┤ĆļĀ©ņØ┤ ņ׳ņØä ņłś ņ׳ļŗż[30]. Ēśäņ×¼Ļ╣īņ¦Ć ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņŚÉ ļīĆĒĢ£ ļČäņ×ÉņĀü ņØ┤ĒĢ┤ļź╝ ļÅĢĻĖ░ ņ£äĒĢ£ ņŚ¼ļ¤¼ ņŚ░ĻĄ¼ļōżņØ┤ ņ¦äĒ¢ēļÉśņŚłņ£╝ļéś, ņĢäņ¦üĻ╣īņ¦Ć ļ¬ģĒÖĢĒĢ£ ĻĖ░ņĀä ĒÖĢņØĖņŚÉļŖö ĒĢ£Ļ│äĻ░Ć ņ׳ļŗż.
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ņäŁņĘ© ļ░Å ĒØĪņłś
ņŚ¼ļ¤¼ ņŚ░ĻĄ¼ņŚÉ ļö░ļź┤ļ®┤ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ĒśłĻ┤ĆņĪ░(stria vascularis)ļź╝ ĒåĄĒĢ┤ ņÜ░ņäĀņĀüņ£╝ļĪ£ ņÖĆņÜ░ļĪ£ ļōżņ¢┤Ļ░ä Ēøä ļé┤ļ”╝ĒöäņŚÉņä£ ņ£Āļ¬©ņäĖĒżļĪ£ ļōżņ¢┤Ļ░äļŗżĻ│Ā ĒĢ£ļŗż[31-33]. ĻĄ¼ļ”¼ ņ£Āņé¼ ņłśņåĪņ▓┤-1 (copper-like transport-1, CTR1) ļ░Å ņ£ĀĻĖ░ ņ¢æņØ┤ņś© ņłśņåĪņ▓┤-2 (organic cation transporter 2, OCT2)ļŖö ĒśłĻ┤ĆņĪ░ņØś ņäĖĒżņŚÉņä£ ļ░£ĒśäļÉśņ¦Ćļ¦ī ļéśņäĀ ņØĖļīĆņŚÉņä£ļŖö ļ░£ĒśäļÉśņ¦Ć ņĢŖļŖö ņĀÉņ£╝ļĪ£ ļ│┤ņĢä, ņØ┤ļōżņØ┤ ĒśłņĢĪ-ļ»ĖļĪ£ ņןļ▓Į(blood labyrinth barrier, BLB)ņØä Ļ░ĆļĪ£ņ¦łļ¤¼ ļé┤ļ”╝Ēöä ļé┤ļĪ£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņłśņåĪĒĢśļŖö ņ×Āņ×¼ņĀüņØĖ ņłśņåĪ ļ®öņ╗żļŗłņ”śņØ╝ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż[34-36].
ņØ╝ļ░śņĀüņ£╝ļĪ£ ņĢĮļ¼╝ņŚÉ ņØśĒĢ£ ņśüĻĄ¼ņĀü ņØ┤ļÅģņä▒ņØ┤ ņØ╝ņ¢┤ļéśĻĖ░ ņ£äĒĢ┤ņä£ļŖö ņØ┤ļÅģņåīĻ░Ć ņ£Āļ¬©ņäĖĒżņŚÉ ļōżņ¢┤Ļ░ĆļŖö Ļ│╝ņĀĢņØ┤ ĒĢäņÜöĒĢ£ļŹ░, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØ┤ ņ£Āļ¬©ņäĖĒżļĪ£ ļōżņ¢┤Ļ░ĆļŖö ņĀĢĒÖĢĒĢ£ ĻĖ░ņĀäņØĆ Ēśäņ×¼Ļ╣īņ¦ĆļŖö ņĢīĻĖ░ ņ¢┤ļĀżņÜ┤ ņāüĒā£ņØ┤ļéś, Hall ļō▒ņØś ņŚ░ĻĄ¼ņŚÉ ļö░ļź┤ļ®┤ ņłśļÅÖņĀüņØĖ ĒÖĢņé░ Ļ│╝ņĀĢņØä ĒåĄĒĢ┤ņä£ļÅä ļ░░ņ¢æļÉ£ ņ£Āļ¬© ņäĖĒżļĪ£ ļōżņ¢┤Ļ░ł ņłś ņ׳ņØīņØ┤ ļ│┤Ļ│ĀļÉśņŚłļŗż[37,38]. ļśÉĒĢ£ ņ£Āļ¬©ņäĖĒżņØś stereocilia ļüØļČĆļČäņŚÉļŖö ļ╣äņäĀĒāØņĀü ņ¢æņØ┤ņś©ņ▒äļäÉņØĖ ļ¦ēĒł¼Ļ│╝ ņ▒äļäÉ ņ£Āņé¼ ļŗ©ļ░▒ņ¦ł(transmembrane channel-like protein-1, TMC1)ņØ┤ ņ׳ņ¢┤ ļ¼╝ņ¦łņØś ĻĖ░Ļ│äņĀü Ļ░£ĒÅÉņŚÉ ņ×æņÜ®ĒĢśĻ▓ī ļÉśļŖöļŹ░, ņĀ£ļĖīļØ╝Ēö╝ņē¼ļź╝ ņØ┤ņÜ®ĒĢ£ Ļ│╝Ļ▒░ ņŚ░ĻĄ¼ņŚÉņä£ļŖö ĻĖ░ļŖźņĀü TMC1 ņ▒äļäÉņØä Ļ░Ćņ¦ä ņ£Āļ¬©ņäĖĒżņØś Ļ▓ĮņÜ░ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņŚÉ Ļ░Ćņן ņĘ©ņĢĮĒĢ©ņØä ĒÖĢņØĖĒĢ©ņ£╝ļĪ£ņŹ©, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņ£Āļ¬©ņäĖĒżļĪ£ ņłśņåĪļÉśļŖö Ļ│╝ņĀĢņŚÉ TMC1 ņ▒äļäÉņØ┤ Ļ┤ĆņŚ¼ĒĢĀ Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż[39-41]. ĒśłĻ┤ĆņĪ░ņäĖĒżņŚÉņä£ ļ░£ĒśäļÉśļŖö CTR1, OCT2 ņłśņåĪņ▓┤ļŖö ņÖĆņÜ░ ņ£Āļ¬©ņäĖĒżņŚÉņä£ļÅä ļ░£ĒśäņØä ĒÖĢņØĖĒĢ©ņ£╝ļĪ£ņŹ©, ņØ┤ļōżņØ┤ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ņ£Āļ¬©ņäĖĒżļĪ£ņØś ĒØĪņłśļÅä ļÅĢļŖö Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż[40,41]. OCT ņ░©ļŗ©ņĀ£ņØĖ ņŗ£ļ®öĒŗ░ļöśņØä ņĀäņŗĀĒł¼ņŚ¼ĒĢśĻ▒░ļéś OCTņØś ņ£ĀņĀäņĀü ĒāłļØĮņØä ņ£Āļ░£ĒĢśļŖö ĒÖ®ņé░ĻĄ¼ļ”¼ļź╝ ĻĄŁņåī Ēł¼ņŚ¼ĒĢ£ ņŚ░ĻĄ¼ņŚÉņä£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØś ņÖäĒÖö ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢśņŚ¼ OCTĻ░Ć ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ņ£Āļ¬©ņäĖĒżļĪ£ņØś ņłśņåĪņŚÉ Ļ┤ĆņŚ¼ĒĢ©ņØä ļÆĘļ░øņ╣©ĒĢśļŖö ĻĘ╝Ļ▒░ļĪ£ ņĀ£ņŗ£ļÉśņŚłņ£╝ļéś, ņĀ£ļĖīļØ╝Ēö╝ņē¼ ņŚ░ĻĄ¼ ļō▒ņŚÉņä£ļŖö ņØ┤ļ¤¼ĒĢ£ ņłśņåĪņ▓┤ņØś ļ░£ĒśäņØ┤ ļ░£Ļ▓¼ļÉśņ¦Ć ņĢŖņĢä, Ēśäņ×¼Ļ╣īņ¦ĆļŖö TMC1 ņ▒äļäÉņØ┤ ņŻ╝ļÉ£ ņłśņåĪ ņŚŁĒĢĀņØä ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż[34,35]. ĒØĪņłśļÉ£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņŚ╝ĒÖöļ¼╝ ņżæ ĒĢśļéś ļśÉļŖö ļæś ļ¬©ļæÉĻ░Ć ļ¼╝ ļČäņ×ÉņŚÉ ņØśĒĢ┤ ļīĆņ▓┤ļÉśņ¢┤ ļé«ņØĆ ļ¦ēĒł¼Ļ│╝ņä▒ņØä Ļ░¢ļŖö ņĢäņ┐ĀņĢäļ│ĄĒĢ®ņ▓┤ļź╝ ņāØņä▒ĒĢśļŖö ņłśĒÖö Ļ│╝ņĀĢņØä Ļ▒░ņ╣£ļŗż[42]. ļśÉĒĢ£ ņ£Āļ¬©ņäĖĒżļĪ£ ļōżņ¢┤Ļ░ä ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņĄ£ņåī 12-18Ļ░£ņøö ļÅÖņĢł ņ£Āņ¦ĆļÉśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Fig. 1) [31].
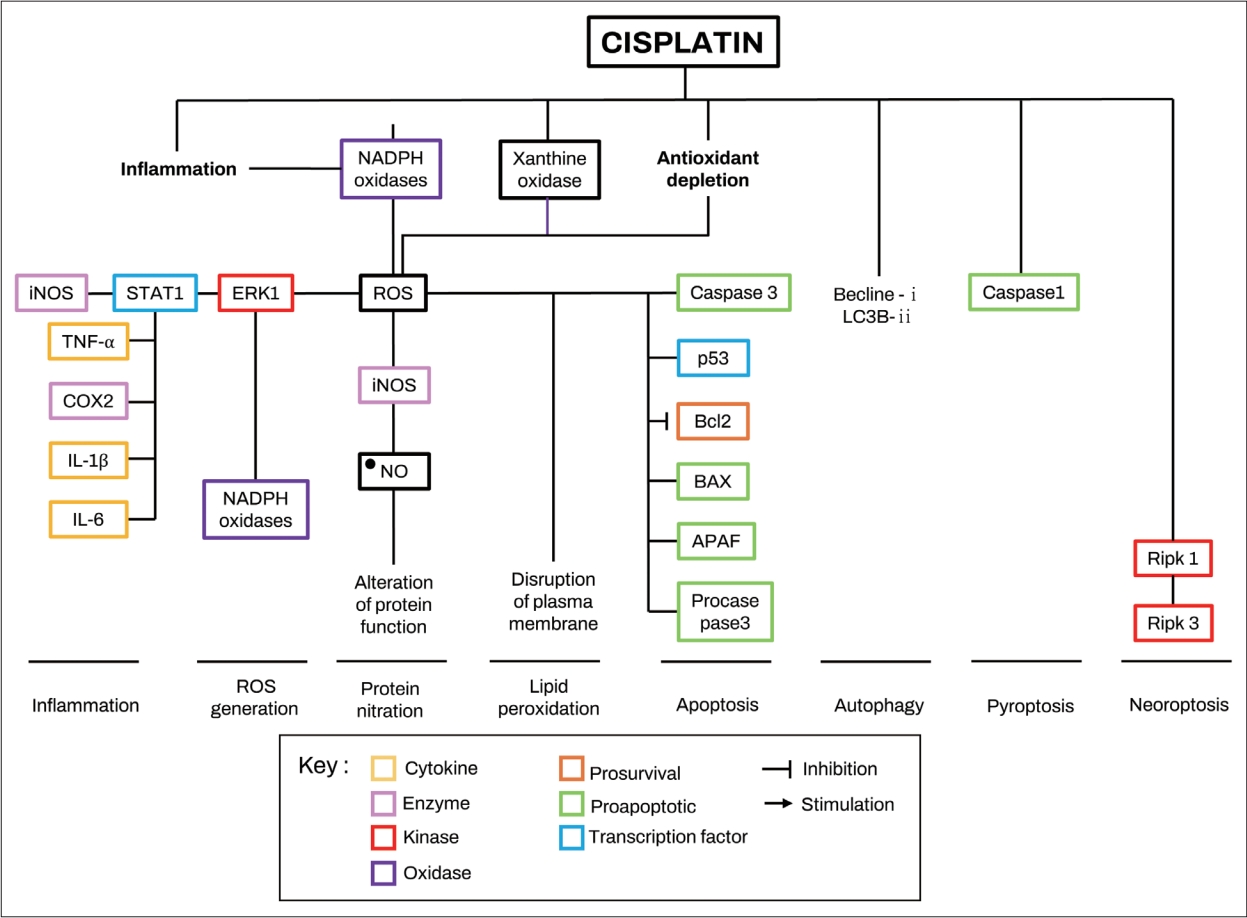
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ£ ņØ┤ļÅģņä▒ņØś ļČäņ×É ĻĖ░ņĀä
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņ£Āļ¬©ņäĖĒżļĪ£ ļōżņ¢┤Ļ░Ćļ®┤ ņäĖĒżņé¼ļ®ĖņØś ņŚ¼ļ¤¼ ļ®öņ╗żļŗłņ”śņØä ņ£Āļ░£ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ļŗż[43,44]. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņØ╝ņ░©ņĀüņ£╝ļĪ£ ņäĖĒżļČäņŚ┤ Ļ│╝ņĀĢņŚÉņä£ DNAņÖĆ Ļ▓░ĒĢ®ĒĢśņŚ¼ ņäĖĒżņ×Éļ®Ėņé¼ļź╝ ņ£Āļ░£ĒĢ£ļŗż. ĻĘĖļ¤¼ļéś ņäĖĒżļČäņŚ┤ņØä ĒĢśņ¦Ć ņĢŖļŖö Ēżņ£ĀļÅÖļ¼╝ņØś ņ£Āļ¬©ņäĖĒżņŚÉļÅä ņäĖĒż ļé┤ Ļ▓ĮļĪ£ņÖĆ ļŗ©ļ░▒ Ļ▓░ĒĢ®ņŚÉ ņśüĒ¢źņØä ņŻ╝ņ¢┤ ņäĖĒż ņĪ░ņĀłņØä ņåÉņāüņŗ£ĒéżĻ│Ā ļŗ©ļ░▒ Ļ▓░ĒĢ®ņŚÉ ņśüĒ¢źņØä ņŻ╝ņ¢┤ ņäĖĒżņé¼ļ®ĖņØä ņ£Āļ░£ĒĢĀ ņłś ņ׳ļŗż(Fig. 2) [44-46].
ņŚ╝ņ”Ø(Inflammation)
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØĆ ņŚ╝ņ”Øļ░śņØæņØś ņ×ÉĻĘ╣Ļ│╝ Ļ┤ĆļĀ©ņØ┤ ņ׳ļŖöļŹ░, ņäĖĒżņÖĖ ņŗĀĒśĖņĪ░ņĀł ĒéżļéśņĢäņĀ£(extracellular signal-regulated kinases, ERK)ļź╝ ĒÖ£ņä▒ĒÖö ĒĢ©ņ£╝ļĪ£ņŹ©, ņóģņ¢æĻ┤┤ņé¼ņØĖņ×É(tumor necrosis factor ╬▒, TNF-╬▒), ņØĖĒä░ļŻ©Ēé©-1b (interleukin 1b, IL-1b), IL-6ņØś ļ░®ņČ£ņØä ņ£ĀļÅäĒĢ£ļŗż[47-49]. ņĮöļź┤Ēŗ░ĻĖ░Ļ┤ĆņŚÉņä£ ĒīīņāØļÉ£ ļ¦łņÜ░ņŖż ņäĖĒż ļØ╝ņØĖņØĖ HEI-OC1 ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņ▓śļ”¼ 1ņŗ£Ļ░ä Ēøä ĻĖ░ņĪ┤ ņäĖĒżņŚÉņä£ ņŚ╝ņ”Ø ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØ┤ ļČäļ╣äļÉśĻ│Ā, ņØ┤ĒøäņŚÉļŖö ņāłļĪ£ņÜ┤ ļŗ©ļ░▒ ĒĢ®ņä▒ņØä ĒåĄĒĢ┤ ņŚ╝ņ”Ø ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØ┤ ļČäļ╣äļÉ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[47]. ņØ┤ļ¤¼ĒĢ£ Ļ▓░Ļ│╝ļŖö ņŚ╝ņ”ØņØ┤ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØś ņ▓½ļ▓łņ¦Ė Ļ│╝ņĀĢņØ╝ Ļ▓āņØ┤ļØ╝ļŖö Ļ░ĆņäżņØä ļÆĘļ░øņ╣©ĒĢ£ļŗż. ļśÉĒĢ£ 4ņØ╝Ļ░ä 4 mg/kgņØś ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ļ│ĄĻ░Ģ ņŻ╝ņé¼ĒĢ£ ļ¦łņÜ░ņŖżņŚÉņä£ļŖö TNF-╬▒, IL-1b ļ░Å IL-6ņØś Ēśłņ▓Ł ņłśņżĆņØ┤ 3-7ļ░░ ņ”ØĻ░ĆĒ¢łņ£╝ļ®░, ņÖĆņÜ░ņŚÉņä£ ļÅÖņØ╝ĒĢ£ ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØś mRNA ļ░Å ļŗ©ļ░▒ ņłśņ╣śĻ░Ć ņ”ØĻ░ĆĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[47]. ĒŖ╣Ē׳, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņ£ĀļÅä TNF-╬▒ ļŗ©ļ░▒ ņłśņ╣śļŖö ņĮöļź┤Ēŗ░ĻĖ░Ļ┤ĆņØś ĒśłĻ┤ĆņĪ░, ļéśņäĀņØĖļīĆ(spiral ligament)ņÖĆ ļéśņäĀņ£żļČĆ(spiral limbus), spiral modiolar veins and lacunaeĻ│╝ ņ£Āļ¬©ņäĖĒżņŚÉņä£ ņ”ØĻ░ĆļÉ©ņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ®░, IL-1bļŖö ņŻ╝ļĪ£ ļéśņäĀ ņØĖļīĆņÖĆ spiral modiolar veins ļ░Å lacunaeņŚÉņä£ ļ░£Ļ▓¼ļÉ£ ļ░śļ®┤, IL-6ļŖö spiral modiolar veins ļ░Å lacunaeņŚÉņä£ļ¦ī ļ░£Ļ▓¼ļÉśņŚłļŗż[47]. ĒĢŁņé░ĒÖöņĀ£ ņĀäņ▓śļ”¼ļź╝ ĒĢ£ HEI-OC1 ņäĖĒżņŚÉņä£ļŖö ņé¼ņØ┤ĒåĀņ╣┤ņØĖ ļČäļ╣äĻ░Ć ņśüĒ¢źņØä ļ░øņ¦Ć ņĢŖņĢśļŖöļŹ░, ņØ┤ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņ£ĀļÅä ņŚ╝ņ”Øļ░śņØæņØ┤ ĒÖ£ņä▒ņé░ņåīņóģ(reactive oxygen species, ROS)ņØś ļ░£ņāØļ│┤ļŗż ņäĀĒ¢ēĒĢśļŖö ļ░śņØæņ×äņØä ņŗ£ņé¼ĒĢ£ļŗż[47]. ļśÉĒĢ£, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ Ēł¼ņŚ¼ ņĀä TNF-╬▒ļź╝ ņżæĒÖöĒĢśĻ▓ī ļÉśļ®┤ HEI-OC1 ņäĖĒżņØś ņāØņĪ┤ļĀźņØ┤ Ē¢źņāüļÉśņŚłĻ│Ā, UB-OC1 ņäĖĒżņŚÉņä£ļŖö ņäĖĒżņ×Éļ®Ėņé¼Ļ░Ć Ļ░ÉņåīĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ®░. TNF-╬▒, IL-1b, IL-6ļź╝ ļÅÖņŗ£ņŚÉ ņżæĒÖöĒĢśļŖö Ļ▓ĮņÜ░ņŚÉļŖö HEI-OC1 ņäĖĒżņŚÉņä£ ROS ņāØņé░ņØ┤ Ļ░ÉņåīĒĢśņśĆļŗż[47]. ņØ┤ļŖö ļé┤ņØ┤ņäĖĒżņŚÉņä£ ņŚ╝ņ”Øņä▒ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ, ņäĖĒżņ×Éļ®Ėņé¼, ROS ņāØņä▒ ņé¼ņØ┤ņŚÉ ļ░ĆņĀæĒĢ£ Ļ┤ĆļĀ©ņØ┤ ņ׳ņØīņØä ņŗ£ņé¼ĒĢ£ļŗż. HEI-OC1 ņäĖĒżļź╝ ĒåĄĒĢ£ ļśÉļŗżļźĖ ņŚ░ĻĄ¼ņŚÉņä£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB)ņØś ņĀäņ£ä ņ”ØĻ░Ć, TNF-╬▒, IL-6ņØś ņ”ØĻ░Ćļź╝ ĒåĄĒĢ┤ ņŚ╝ņ”Øļ░śņØæņØä ņ”ØĒÅŁņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłļŗż[50].
ņĀäņŚ╝ņ”Øņä▒ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ(proinflammatory cytokine)ņØś ĒĢ®ņä▒ņØĆ ņŗĀĒśĖļ│ĆĒÖśĻĖ░ņÖĆ ņĀäņé¼ ĒÖ£ņä▒ņØĖņ×É(signal transducer and activator of transcription 1, STAT1)ņØś ĒÖ£ņä▒ņŚÉ ņØśĒĢ┤ ņ┤ēņ¦äļÉśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖöļŹ░, STAT6 (STAT6-/-)Ļ░Ć ļ╣äĒÖ£ņä▒ĒÖöļÉ£ ļ¦łņÜ░ņŖżņØś ņÖĆņÜ░ņäĖĒżņŚÉņä£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ TNF-╬▒, IL-1b ļ░Å IL-6ņØś Ēśłņ▓ŁņłśņżĆ ļ░Å mRNA ļ░£Ēśä, NK-kB mRNAļź╝ ņ”ØĻ░Ćņŗ£Ēéżņ¦Ć ņĢŖņĢśņ£╝ļ®░, ņäĖĒżņ×Éļ®Ėņé¼ ļśÉĒĢ£ ņ£ĀļÅäĒĢśņ¦Ć ņĢŖņĢä, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØ┤ ļ¦żĻ░£ĒĢśļŖö ņĀäņŚ╝ņ”Øļ░śņØæņŚÉņä£ STAT6Ļ░Ć Ļ┤ĆņŚ¼ĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[50,51]. UB-OC1 ņäĖĒż ņŚ░ĻĄ¼ņŚÉņä£ ĒÖ£ņä▒ĒśĢ STAT1ņØĆ ņŚ╝ņ”Ø ļŗ©ļ░▒ņØĖ cyclooxygenase 2 (COX2), TNF-╬▒, ņ£ĀļÅäņä▒ ņé░ĒÖöņ¦łņåī ĒĢ®ņä▒ ĒÜ©ņåī(inducible nitric oxide synthase, iNOS), ļ®┤ņŚŁņäĖĒżņ╣©ņ£żņØś ļ¦łņ╗żļĪ£ ņĢīļĀżņ¦ä CD14 ļ░Å CD45ņØś ļ░£ĒśäņØä ĒåĄĒĢ┤ ņŚ╝ņ”Øļ░śņØæņØä ņ┤ēņ¦äĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśļ®░, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņŻ╝ņ×ģĒĢ£ ņźÉņØś ĒśłĻ┤ĆņĪ░, ļéśņäĀņØĖļīĆ, ļéśņäĀņ£żļČĆ, ļéśņäĀņŗĀĻ▓ĮņĀł ļ░Å ņÖĖņ£Āļ¬©ņäĖĒżņŚÉņä£ STAT1ņŚÉ ņØśĒĢ┤ ļÅÖņØ╝ĒĢ£ ņŚ╝ņ”Ø ļŗ©ļ░▒Ļ│╝ ļ®┤ņŚŁ ļ¦łņ╗żņØś ņāüņŖ╣ņØ┤ ĒÖĢņØĖļÉśņŚłļŗż[52]. ņØ┤ļ¤¼ĒĢ£ Ļ▓░Ļ│╝ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØ┤ ņÖĆņÜ░ņäĖĒżņŚÉņä£ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ ļ░®ņČ£ņŚÉ ņ¦üņĀæ ņśüĒ¢źņØä ļ»Ėņ╣śĻ│Ā STAT ļŗ©ļ░▒Ļ│╝ NF-kB ĒÖ£ņä▒ņØä Ļ░ĢĒÖöĒĢśņŚ¼ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ ņāØņä▒ņØä ņ×ÉĻĘ╣ĒĢ©ņ£╝ļĪ£ņŹ© ņŚ╝ņ”ØņØä ņ£ĀļÅäĒĢ£ļŗżļŖö Ļ░ĆņäżņØä ļÆĘļ░øņ╣©ĒĢ£ļŗż[52].
ĒÖ£ņä▒ņé░ņåī ņāØņä▒ ļ░Å ņŗĀĒśĖ Ļ▓ĮļĪ£(ROS Generation and Signaling Pathways)
ņÖĆņÜ░ļŖö ļīĆņé¼ ĒÖ£ļÅÖņØ┤ ĒÖ£ļ░£ĒĢ£ ĻĖ░Ļ┤Ćņ£╝ļĪ£ņä£, ņÖĆņÜ░ņØś ĒĢŁņé░ĒÖö ņŗ£ņŖżĒģ£ņŚÉļŖö ĻĖĆļŻ©ĒāĆņ╣śņś© ĒŹ╝ņśźņŗ£ļŗżņĀ£, ĻĖĆļŻ©ĒāĆĒŗ░ņś© ĒÖśņøÉĒÜ©ņåī, ĻĖĆļŻ©ĒāĆĒŗ░ņś©-s-ĒŖĖļ×£ņŖżĒŹ╝ļØ╝ņĀ£, ņŖłĒŹ╝ņśźņé¼ņØ┤ļō£ ļööņŖżļ«żĒāĆņĀ£, ņ╣┤ĒāłļØ╝ņĀ£ ļ░Å Ļ░Éļ¦ł-ĻĖĆļŻ©ĒāĆļ░Ć ņŗ£ņŖżĒģīņØĖ ĒĢ®ņä▒ĒÜ©ņåīņÖĆ Ļ░ÖņØĆ ĒÜ©ņåīĻ░Ć ĒżĒĢ©ļÉśļ®░, ĻĘĀĒśĢņĀüņØĖ ĒĢŁņé░ĒÖö ņŗ£ņŖżĒģ£ņØĆ ņÖĆņÜ░ņØś ņĀĢņāüņĀüņØĖ ņāØļ”¼ņÖĆ ņ▓ŁļĀź ņ£Āņ¦ĆņŚÉ ĒĢäņłśņĀüņØĖ ņŚŁĒĢĀņØä ĒĢ£ļŗż. ņÖĆņÜ░ļŖö ņĀĆņé░ņåīņ”ØĻ│╝ ĒŚłĒśł-ņ×¼Ļ┤ĆļźśņŚÉ ļ¦żņÜ░ ļ»╝Ļ░ÉĒĢ£ ĻĖ░Ļ┤Ćņ£╝ļĪ£ Ļ│╝ļÅäĒĢ£ ņåīņØīĻ│╝ ņØ┤ļÅģņä▒ ļ¼╝ņ¦łņØĆ ņÖĆņÜ░ņØś ņé░ĒÖöņŖżĒŖĖļĀłņŖżļź╝ ņ”ØĻ░Ćņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[29]. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØĆ ņÖĆņÜ░ņØś ĒŖ╣ņØ┤ņĀü ĒÖ£ņä▒ņé░ĒśĖ ņ”ØĻ░Ć ĒÜ©ņåīņØĖ NADPH oxidase 3 (NOX3), Ēü¼ņé░Ēŗ┤ ņé░ĒÖöĒÜ©ņåī(xanthin oxidase) ļ░Å ļé┤ņØĖņä▒ ĒĢŁņé░ĒÖö ĒÜ©ņåī ņŗ£ņŖżĒģ£ņØś Ļ░Éņåīļź╝ ĒåĄĒĢ┤ ROSļź╝ ņ”ØĻ░Ćņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ļŗż[7,53-56]. ņÖĆņÜ░ņäĖĒżņŚÉņä£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ĒØĪņłśļŖö ņĀäņé¼ņØĖņ×É, ņŗĀĒśĖļ│ĆĒÖöņØĖņ×É ļ░Å STAT1ņØä ņ×ÉĻĘ╣ĒĢśņŚ¼ TRPV1Ļ│╝ NOX3 Ļ▓ĮļĪ£ļź╝ ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ĒÖ£ņä▒ņé░ņåīļź╝ ļÅģņä▒ ņłśņżĆĻ╣īņ¦Ć ļ░£ņāØņŗ£ņ╝£ ņäĖĒż ņé¼ļ®ĖņØä ņ£Āļ░£ĒĢśļ®░, ņØ┤ļŖö ņ£Āļ¬©ņäĖĒżļ┐Éļ¦ī ņĢäļŗłļØ╝ ņ£Āļ¬©ņäĖĒż ĒĢŁņāüņä▒ ņ£Āņ¦ĆņŚÉ ņżæņÜöĒĢ£ ļ╣äĻ░ÉĻ░ü ņÖĆņÜ░ņäĖĒż ļ░Å ļé┤ļ”╝Ēöä ņĀäņ£äļź╝ ņ£Āļ░£ĒĢśļŖö ĒśłĻ┤ĆņĪ░ņØś ņżæĻ░äņäĖĒż(intermediate cell) ļ░Å ļ│ĆņŚ░ņäĖĒż(marginal cell)ļÅä ĒżĒĢ©ļÉ£ļŗż[57-59].
ROS ņāØņä▒ņØĆ ļŗżņ¢æĒĢ£ ĒĢŁņé░ĒÖöņĀ£ ĒÜ©Ļ│╝ņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņ£Āļ░£ ļé£ņ▓ŁņØś ĻĖ░ļ│Ė ĻĖ░ņĀäņ£╝ļĪ£ ņĀ£ņĢłļÉ£ļŗż[60,61]. ņÖĆņÜ░ņäĖĒżņŚÉņä£ ROS ņāØņä▒ņØä ņ£Āļ░£ĒĢśļŖö ņŻ╝ņÜö Ļ│╝ņĀĢņØĆ NADPH ņé░ĒÖöĒÜ©ņåī ņåīļŗ©ņ£äņØś ņāüĒ¢źņĪ░ņĀłļĪ£ ĒÖĢņØĖļÉśņŚłņ£╝ļ®░[62-64], Ļ░ĢĒÖöļÉ£ NADPH ņé░ĒÖöĒÜ©ņåī ĒÖ£ņä▒ņØä ĒåĄĒĢ┤ ņāØņä▒ļÉ£ ROSļŖö NADPH ņé░ĒÖöĒÜ©ņåī ņåīļŗ©ņ£äņØś ĒĢ®ņä▒ņØä ļŗżņŗ£ ņ×ÉĻĘ╣ĒĢśņŚ¼ ņ×ÉĻ░Ć ņ┤ēļ¦ż ļŻ©ĒöäņŚÉņä£ ROS ņāØņé░ņØä ļŹöņÜ▒ ņ”ØĻ░Ćņŗ£Ēé©ļŗż[64]. HEI-OC1 ņäĖĒżņŚ░ĻĄ¼ņŚÉņä£ 6ņŗ£Ļ░ä ļÅÖņĢł 20 mM ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ļģĖņČ£ļÉ£ TNF-╬▒, IL-1b ļ░Å IL-6ļŖö NADPH ņé░ĒÖöĒÜ©ņåī ĒÖ£ņä▒ņØä 20%-50% Ļ░Éņåīņŗ£ņ╝░ņ£╝ļ®░, ļśÉļŗżļźĖ ROS ņāØņä▒ ĒÜ©ņåīņØĖ xanthine oxidaseņØś ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£Ēé┤ņ£╝ļĪ£ņŹ© ROS ņāØņé░ņØä Ē¢źņāüņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņŚłļŗż[64,65]. ņĄ£ĻĘ╝ ņŚ░ĻĄ¼ņŚÉņä£ļŖö 18ņŗ£Ļ░ä ļÅÖņĢł 50 mMņØś ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņ£╝ļĪ£ ņ▓śļ”¼ĒĢ£ HEIOC1 ņäĖĒżņŚÉņä£ ļ│┤ņĪ░ņØĖņ×É ņé¼ņØ┤Ēü┤ļ”░ A (cofactor cyclin A)ņØś ņāüĒ¢źņĪ░ņĀłņØä ĒåĄĒĢ┤ ņé¼ņØ┤Ēü┤ļ”░ ņØśņĪ┤ņä▒ ĒéżļéśņĀ£ 2 (cyclin-dependent kinase 2, CDK2)ĒÖ£ņä▒ņØ┤ 2ļ░░ ņ”ØĻ░ĆĒĢśļŖö Ļ▓āņØ┤ ļ░ØĒśĆņĪīņ£╝ļ®░, CDK2 ņ¢ĄņĀ£ņĀ£ļĪ£ ņ▓śļ”¼ĒĢ£ ļ¦łņÜ░ņŖż explantņÖĆ CDK2-knockout cochlear ex- plantņŚÉņä£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņ£ĀļÅä ROS ņāØņä▒ ļ░Å ņäĖĒż ņ×Éļ®Ėņé¼Ļ░Ć ņ£ĀņØśĒĢśĻ▓ī Ļ░ÉņåīĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[66]. ĻĘĖļ¤¼ļéś ņĢäņ¦üĻ╣īņ¦Ć ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØ┤ ņé¼ņØ┤Ēü┤ļ”░ A ļ░Å CDK2 ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£ĒéżļŖö ĻĖ░ņĀäņØĆ ļ¬ģĒÖĢĒ׳ ļ░ØĒśĆņĀĖ ņ׳ņ¦Ć ņĢŖļŗż.
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ┤ ņ”ØĻ░ĆļÉ£ ROSļŖö extracellular signalregulated kinase 1 (ERK1)/STAT1 Ļ▓ĮļĪ£ļź╝ ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ņŚ╝ņ”ØĻ│╝ ņäĖĒżņ×Éļ®Ėņé¼ņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ļŗ©ļ░▒ņØĖ COX2, TNF-╬▒, iNOS, p53, caspase 3 ļ░Å B-cell lymphoma 2 (Bcl-2)-like protein 4 (BAX)ņØś ļŹö Ļ░ĢĒĢ£ ļ░£ĒśäņØä ņ£Āļ░£ĒĢ£ļŗż[67]. ļśÉĒĢ£ ROSļŖö DNA ņåÉņāüņØä ņØ╝ņ£╝ņ╝£ DNA ļ│ĄĻĄ¼ļŗ©ļ░▒ņ¦ł Ļ│╝ĒÖ£ņä▒ĒÖöļź╝ ņ£Āļ░£ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖöļŹ░[68], 16 mg/kgņØś ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņŻ╝ņ×ģĒĢ£ ļ¦łņÜ░ņŖżņØś ņÖĆņÜ░ņŚÉņä£ 4ņØ╝ Ēøä ĒÅ┤ļ”¼ ADP-ļ”¼ļ│┤ņŖż ĒŖĖļ×£ņŖżĒŹ╝ļØ╝ņĀ£ 1(poly-ADP-ribose transferase 1, PARP-1) ĒÖ£ņä▒ņØ┤ 100% ņ”ØĻ░ĆĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[69]. PARP-1ņØĆ DNA ļ│ĄĻĄ¼ļŗ©ļ░▒ņ¦łņØś ņØ┤ļÅÖņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ĒÜ©ņåīņØ┤ļ®░, ņØ┤ņØś ņ¢ĄņĀ£ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ£ ņäĖĒż ņé¼ļ®Ė Ļ░ÉņåīņÖĆ Ļ┤ĆļĀ©ņØ┤ ņ׳ļŖö Ļ▓āņØä ņØśļ»ĖĒĢ£ļŗż[69]. ļśÉĒĢ£, 20 mM ļåŹļÅäņØś ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņ£╝ļĪ£ ņ▓śļ”¼ĒĢ£ HEI-OC1 ņäĖĒżņŚÉņä£ļŖö NAD+/NADH ļ╣äņ£©ņØ┤ ņŗ£Ļ░äņŚÉ ļö░ļØ╝ Ļ░Éņåī(12ņŗ£Ļ░ä Ēøä ļīĆņĪ░ĻĄ░ ļīĆļ╣ä -30% ļ░Å 24ņŗ£Ļ░ä Ēøä -55%)ĒĢśļŖö Ļ▓āņØ┤ ĒÖĢņØĖļÉśņ¢┤, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ļģĖņČ£ ņ▓ŁĻ░üņäĖĒżĻ░Ć DNA ļ│ĄĻĄ¼ ļ®öņ╗żļŗłņ”śņØä ĒÖ£ņä▒ĒÖöĒĢśņ¦Ćļ¦ī DNA ņåÉņāüņØä ņśłļ░®ĒĢĀ ņłś ņŚåņØīņØä ņØśļ»ĖĒĢ£ļŗż[69].
ļŗżļźĖ ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņŻ╝ņ×ģĒĢ£ ņźÉņŚÉņä£ ROS ņ£ĀļÅä ņ¦Ćņ¦łĻ│╝ņé░ĒÖö(llipi peroxidation)ļź╝ ĒåĄĒĢ£ ĒĢŁņé░ĒÖö ļ░®ņ¢┤ņŗ£ņŖżĒģ£ņØś Ļ▓░ĒĢŹ ļ░Å ņäĖĒżņåÉņāüņØä ĒÖĢņØĖĒĢśņśĆĻ│Ā, UB-OC1 ņäĖĒżņŚÉņä£ļŖö ņÖĆņÜ░ļŗ©ļ░▒ņ¦łņØś nitrationņØä ĒåĄĒĢ£ ņåÉņāüņØä ĒÖĢņØĖĒĢśņśĆņ£╝ļ®░, ļæÉ ĻĖ░ņĀä ļ¬©ļæÉ ĻČüĻĘ╣ņĀüņ£╝ļĪ£ ņäĖĒżņ×Éļ®Ėņé¼ļĪ£ ņ¦äĒ¢ēļÉ©ņØ┤ ļ░ØĒśĆņĪīļŗż[70-72].
ņóģĒĢ®ĒĢśļ®┤, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ NADPH ņé░ĒÖöĒÜ©ņåīņÖĆ xanthine oxidase ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£ĒéżĻ│Ā ĒĢŁņé░ĒÖö ļ░®ņ¢┤ņŗ£ņŖżĒģ£ņØä ļ░®ĒĢ┤ĒĢśļ®░ ņŚ╝ņ”Øņä▒ ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØś ļČäļ╣äļź╝ ņ£ĀļÅäĒĢ©ņ£╝ļĪ£ņŹ© ņÖĆņÜ░ņäĖĒżņŚÉņä£ ROS ņāØņä▒ņØä ņ”ØĻ░Ćņŗ£Ēé©ļŗżļŖö Ļ▓āņØä ļ│┤ņŚ¼ņżĆļŗż. ļśÉĒĢ£ ņ”ØĻ░ĆļÉ£ ROSļŖö ņ¦Ćņ¦ł Ļ│╝ņé░ĒÖö, ļŗ©ļ░▒ņ¦ł ņ¦łĒÖö ļ░Å DNA ņåÉņāüņØä ņ┤ēņ¦äĒĢśĻ│Ā NAD+/NADH ļ╣äņ£©ņØä Ļ░Éņåīņŗ£ņ╝£ Ļ▓░ĻĄŁ ņ▓ŁĻ░ü ņäĖĒż ņé¼ļ®ĖņØä ņ┤łļלĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤ņØĖļŗż[70-72].
ņäĖĒż Ļ│Āņé¼(Cell Death)
ņŚ╝ņ”Ø, ROS ņāØņä▒, ER ņŖżĒŖĖļĀłņŖż, DNA ņåÉņāüņØĆ ņäĖĒżņ×Éļ®Ėņé¼ Ļ▓ĮļĪ£ļź╝ ĒÖ£ņä▒ĒÖöĒĢśļŖö ļŹ░ Ļ┤ĆņŚ¼ĒĢ£ļŗż. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņ▓śļ”¼ĒĢ£ explantņŚÉņä£ļŖö ņÖĖņØĖņä▒ ņäĖĒżņ×Éļ®Ėņé¼ Ļ▓ĮļĪ£ņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö Fas-associated protein with death domain (FADD)ņØĆ ļ░£ĒśäĒĢśņ¦Ć ņĢŖļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉśņŚłņ£╝ļéś[73], ļÅÖņØ╝ĒĢ£ Ļ▓ĮļĪ£ņØś ĻĄ¼ņä▒ņÜöņåī ņżæ ĒĢśļéśņØĖ caspase 8ņØĆ ļ░£ĒśäļÉśļŖö ļ¬©ņł£ņĀÉņØ┤ ņ׳ņ¢┤, ņŗ£ņŖżĒöīļØ╝Ēŗ┤Ļ│╝ ņÖĖņØĖņä▒ ņäĖĒżņ×Éļ®Ėņé¼ņŚÉ ļīĆĒĢ£ ĻĖ░ņĀäņØĆ ņČöĻ░Ć ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢ£ ņāüĒā£ņØ┤ļŗż[74]. ņÖĖņØĖņä▒ ņäĖĒżņ×Éļ®Ėņé¼ņÖĆļŖö ļīĆņĪ░ņĀüņ£╝ļĪ£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ļé┤ņØĖņä▒ ņäĖĒżņ×Éļ®Ėņé¼ņÖĆ Ļ┤ĆļĀ©ļÉ£ proapoptotic transcription factor forkhead box O3 (FOXO3a) ļ░Å BAX ļŗ©ļ░▒ņ¦łņØś ļ░£Ēśä ņłśņżĆņØä ņ”ØĻ░Ćņŗ£ĒéżĻ│Ā BAX ļŗ©ļ░▒ņ¦łņØś ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ļ¦ēņ£╝ļĪ£ņØś ņĀäņ£äļź╝ ņ┤ēņ¦äĒĢ©ņ£╝ļĪ£ņŹ©, ļé┤ņØĖņä▒ ņäĖĒżņ×Éļ®Ėņé¼ Ļ▓ĮļĪ£ļź╝ ĒÖ£ņä▒ĒÖöĒĢ£ļŗżļŖö ņĀÉņØĆ ņŚ¼ļ¤¼ ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ĒÖĢļ”ĮļÉ£ ņé¼ņŗżņØ┤ļŗż[73-75]. ļśÉĒĢ£ ņØ┤ļŖö Bcl-2 ļŗ©ļ░▒ņ¦łņØś Ļ░Éņåī, ļ»ĖĒåĀņĮśļō£ļ”¼ņĢä ņÖĖļ¦ēņØś Ēł¼Ļ│╝, ņé¼ņØ┤ĒåĀĒü¼ļĪ¼ cņØś ļ░®ņČ£, ņäĖĒżņ×Éļ®Ėņé¼-ņ£ĀļÅä ņØĖņ×É(apoptosis-inducing factor, AIF)ņØś ļŹö ļåÆņØĆ ļ░£ĒśäņØä ņ£Āļ░£ĒĢśņŚ¼, ņØ┤ļōżņØś ļ░®ņČ£ ļ░Å ĒĢĄņĀäņ£äļź╝ ņ┤ēņ¦äĒĢ£ļŗż[74-77]. ņØ┤ļŖö ļśÉĒĢ£ ņäĖĒżņé¼ļ®Ė ĒöäļĪ£ĒģīņĢäņĀ£ ĒÖ£ņä▒ĒÖöņØĖņ×É 1 (apoptotic protease activating factor 1, APAF-1)ņØś mRNA ļ░Å ļŗ©ļ░▒ņØä ņ”ØĻ░Ćņŗ£ĒéżĻ│Ā, caspase 9 ļ░Å 3ņØś ļ░£Ēśä ļ░Å ĒÖ£ņä▒ņØä ņ”ØĻ░Ćņŗ£Ēéżļ®░[73,76,77], NF-kB ĒĢĄņ£╝ļĪ£ņØś ņĀäņ£äļź╝ ņ┤ēņ¦äĒĢśĻ│Ā, ņĀäņé¼ņØĖņ×ÉņØś ņäĖĒżņ×Éļ®Ėņé¼ ņŚŁĒĢĀņØä ļÅĢļŖöļŗż[77].
ļŗżņ¢æĒĢ£ ņ▓ŁĻ░üņäĖĒż ļ¬©ļŹĖņŚ░ĻĄ¼ņŚÉņä£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ĒÖ£ņä▒ĒśĢ caspase 1ņØä ņ”ØĻ░Ćņŗ£Ēé┤ņØä ĒÖĢņØĖĒĢśņśĆĻ│Ā, ņØ┤ ĒÜ©ņåīļŖö ņŚ╝ņ”Øņä▒ ņé¼ņØ┤ĒåĀņ╣┤ņØĖ IL-1b ļ░Å IL-18ņØś ņä▒ņłÖ ļ░Å ļČäļ╣äļź╝ ņ┤ēņ¦äĒĢśļŖö ļ│ĄĒĢ®ņ▓┤ņØĖ ņØĖĒöīļØ╝ļ¦łņå£ņØä ĒåĄĒĢ┤ ņäĖĒżņé¼ļ®ĖņØä ņ£ĀļÅäĒĢśļŖö ĒīīņØ┤ļĪŁĒåĀņŗ£ņŖż(pyroptosis) ņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņŚÉ ņØśĒĢ£ ņäĖĒżņé¼ļ®ĖņØ┤ pyroptosisņÖĆļÅä Ļ┤ĆļĀ©ļÉ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[78,79].
ļśÉĒĢ£ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņ×ÉĻ░ĆĒżņŗØ(autophagy)ņØś ļ¦łņ╗żļĪ£ ņל ņĢīļĀżņ¦ä Beclin-1, ļ»ĖņäĖņåīĻ┤Ć Ļ┤ĆļĀ© ļŗ©ļ░▒ņ¦ł ņ▓┤ņØĖ(microtubule-associated protein light chain 3, LC3B-II), ļē┤Ēü┤ļĀłņśżĒŗ░ļō£ Ļ▓░ĒĢ® ņś¼ļ”¼Ļ│Āļ©Ė ļÅäļ®öņØĖ, ļźśņŗĀ ĒĢ©ņ£Ā ļ░śļ│Ąņ▓┤ X1 (leucine-rich repeat-containing X1, NLRX1)ņØś ļ░£ĒśäņØä ņ”ØĻ░Ćņŗ£ņ╝£, ņäĖĒżņ¦łņŚÉ ņ×ÉĻ░ĆĒżņŗØņåīņ▓┤ļź╝ ņČĢņĀüĒĢśņŚ¼ ņäĖĒżņ×Éļ®Ėņé¼ņÖĆļŖö ļÅģļ”ĮņĀüņ£╝ļĪ£ ņ×ÉĻ░ĆĒżņŗØņäĖĒżļź╝ ņé¼ļ®Ėņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ļ│┤ņØĖļŗż[80-82].
ņĄ£ĻĘ╝ ņŚ░ĻĄ¼ņŚÉ ļö░ļź┤ļ®┤ Ļ┤┤ņé¼(necroptosis: a caspase-independent pathway of programmed necrosis)ļÅä ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒Ļ│╝ Ļ┤ĆļĀ©ļÉĀ ņłś ņ׳ņØīņØ┤ ņĢīļĀżņĪīļŖöļŹ░, 16 mg/kg ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ļ│ĄĻ░Ģ ļé┤ ņŻ╝ņ×ģĒĢ£ ņźÉņØś ņÖĆņÜ░ņŚÉņä£ ņłśņÜ®ņ▓┤-ņāüĒśĖņ×æņÜ® ņäĖļ”░/ĒŖĖļĀłņśżļŗī-ļŗ©ļ░▒ņ¦ł ĒéżļéśņĀ£1 (receptor-interacting serine/threonine-protein kinase 1, RIPK1)ņØś ļ░£ĒśäņØ┤ 100% ņ”ØĻ░ĆļÉ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż[83]. RIPK1ņØĆ RIPK3Ļ│╝ ĒĢ©Ļ╗ś ripoptosomeņØä ĒśĢņä▒ĒĢśņŚ¼ Ļ┤┤ņé¼ļź╝ ņ£Āļ░£ĒĢśļŖö ļ¼╝ņ¦łļĪ£[84], ļ®┤ņŚŁņĪ░ņ¦üĒÖöĒĢÖ ļČäņäØ ņŚ░ĻĄ¼ņŚÉņä£ļŖö ļéśņäĀņŗĀĻ▓ĮņĀł, ļé┤ņ£Āļ¬©ņäĖĒżņÖĆ ņÖĖņÜ░ļ¬©ņäĖĒżņØś ļē┤ļ¤░ņŚÉņä£ RIPK3 ļŗ©ļ░▒ ļ░£ĒśäņØ┤ ņ”ØĻ░ĆļÉ©ņØä ĒÖĢņØĖĒĢśņśĆĻ│Ā[83], ļśÉļŗżļźĖ ņŚ░ĻĄ¼ņŚÉņä£ļŖö RIPK1ņØä ņĢĮļ”¼ĒĢÖņĀüņ£╝ļĪ£ ņ¢ĄņĀ£ĒĢśņŚ¼ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ Ļ┤ĆļĀ© ņ▓ŁļĀź ņāüņŗżļĪ£ļČĆĒä░ ļ¦łņÜ░ņŖżļź╝ ļ│┤ĒśĖĒĢśņŚ¼ ABR ņŚŁņ╣śļź╝ 30 dB Ē¢źņāüņŗ£ĒéżļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż[85]. RIPK3-knock out ļ¦łņÜ░ņŖżņÖĆ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņ£╝ļĪ£ ņ▓śļ”¼ļÉ£ Ripk3/caspase 8 double knockout ļ¦łņÜ░ņŖżļŖö ņ£Āļ¬©ņäĖĒż ņāØņĪ┤Ļ│╝ ņ▓ŁļĀź ņŚŁņ╣ś(10 dB)ņŚÉņä£ ņāüļŗ╣ĒĢ£ Ļ░£ņäĀņØä ļ│┤ņØ┤ĻĖ░ļÅä ĒĢśņśĆļŗż[85]. ņØ╝ļČĆņŚÉņä£ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØ┤ caspase 8ņŚÉ ņØśĒĢ┤ ļ¦żĻ░£ļÉśļŖö ņäĖĒżņ×Éļ®Ėņé¼ļ│┤ļŗż ņÖĆņÜ░ņäĖĒżņØś Ļ┤┤ņé¼ ņ┤ēņ¦äņØ┤ ņØ┤ļÅģņä▒ņŚÉ ļŹö ĻĖ░ņŚ¼ĒĢĀ Ļ▓āņØ┤ļØ╝Ļ│Ā ņŻ╝ņןĒĢśĻĖ░ļÅä ĒĢśļéś, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņŚÉņä£ņØś Ļ┤┤ņé¼ ĻĖ░ņĀäņØś ņŚŁĒĢĀņØĆ ņČöĻ░ĆņĀüņØĖ ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢ£ ņāüĒā£ņØ┤ļŗż.
ņØ┤ļÅģņä▒ ņĢģĒÖö ņ£äĒŚśņØä ļåÆņØ┤ļŖö ņÜöņØĖ
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØä Ē¢źņāüņŗ£ĒéżļŖö ņ£äĒŚśņÜöņåīņŚÉļŖö ņŗĀļČĆņĀä ļ░Å Ēāłņłś, ņśüņ¢æ ņŗżņĪ░ ļō▒ņØ┤ ņ׳ņ£╝ļ®░, ĒśłņĢĪ-ļćī ņןļ▓Į(blood-brain barrier) ņØä ĒīīĻ┤┤ĒĢśļŖö ĒöäļĪ£ĒåĀņĮ£ņØś ņé¼ņÜ®ņØĆ ĒśłņĢĪ-ļ»ĖļĪ£ ņןļ▓Į(BLB) ļśÉĒĢ£ ĒīīĻ┤┤ĒĢ©ņ£╝ļĪ£ņŹ© ņØ┤ļÅģņä▒ņØś ņ£äĒŚśņØä ļåÆņØ┤ļŖö Ļ▓āņ£╝ļĪ£ ļ│┤Ļ│ĀļÉ£ļŗż[86,87]. ņ¦Ćņ¦łļŗżļŗ╣ļźś(lipopolysaccharide, LPS)ļĪ£ ņ£ĀļÅäļÉ£ Ēī©Ēśłņ”Ø ļÅÖļ¼╝ļ¬©ļŹĖņŚÉņä£ ņØ┤ļÅģņä▒ņØś ņĢģĒÖöĻ░Ć ļ│┤Ļ│ĀļÉ£ ļ░ö ņ׳ļŖöļŹ░[88], LPSļŖö ņØ╝ļ░śņĀüņ£╝ļĪ£ toll-like receptor 4 (TLR4)ņŚÉ Ļ▓░ĒĢ®ĒĢśņŚ¼ ļé┤ņØĖņä▒ ņåÉņāü Ļ┤ĆļĀ© ļČäņ×ÉĒī©Ēä┤ņØä ņØĖņŗØĒĢśĻ│Ā ņŚ╝ņ”Øļ░śņØæņØä ņ£ĀļÅäĒĢśļŖö ļ¼╝ņ¦łļĪ£ņä£, ņŚ╝ņ”ØĻ│╝ņĀĢņŚÉņä£ ņØ┤ļÅģņä▒ņØś ņĢģĒÖöļź╝ ņØ╝ņ£╝Ēé© Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż[89]. ļśÉĒĢ£, ņ£äņןĻ┤Ć ļ░®ņé¼ņäĀņ╣śļŻīļź╝ ĒĢ©Ļ╗ś ĒĢśļŖö Ļ▓ĮņÜ░ ļ░®ņé¼ņäĀ ņĪ░ņé¼ļŖö Ļ│ĄņāØļ░ĢĒģīļ”¼ņĢäļź╝ ļ░Ģļ®ĖĒĢśĻ│Ā, LPSņØś ĒśłņżæļåŹļÅäļź╝ ļåÆņ×äņ£╝ļĪ£ņŹ© LPS ņ£ĀļÅä ņŚ╝ņ”Øļ░śņØæņØ┤ ļ░£ņāØĒĢśĻĖ░ļÅä ĒĢ┤, ļæÉĻ▓ĮļČĆ ļ░®ņé¼ņäĀ ļ│æņÜ® ņ╣śļŻī ņÖĖņŚÉļÅä ņØ┤ļÅģņä▒ņØś ņ£äĒŚśņØä ņ”ØĻ░Ćņŗ£Ēé¼ ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż[90].
ĒŖ╣ņĀĢ ņ£ĀņĀäņĀü ņåīņØĖ ļśÉĒĢ£ Ļ░£ņØĖņØś ņØ┤ļÅģņä▒ Ļ░Éņłśņä▒ņŚÉ ņśüĒ¢źņØä ņŻ╝ļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŖöļŹ░, ERCC2ņÖĆ XPCņ▓śļ¤╝ DNA ļ│ĄĻĄ¼ ĒÜ©ņåīņŚÉ Ļ┤ĆĒĢ£ ņ£ĀņĀäņ×ÉņØś ļ│ĆņØ┤ ļśÉļŖö ĒĢŁņé░ĒÖö ĒÜ©ņåīļéś, ņĢĮļ¼╝ ņ£ĀņČ£ ļśÉļŖö ņäĖĒżļ¦ē ĒÄīĒöäņÖĆ Ļ┤ĆļĀ©ļÉ£ ABCC3, ACYP2 , COMT, TPMTņØś ļ│ĆņØ┤Ļ░Ć ņØ┤ņŚÉ ĒżĒĢ©ļÉ£ļŗż(Table 1) [91,92]. ĻĘĖļ¤¼ļéś ņĄ£ĻĘ╝ņŚÉļŖö ņØ┤ļ¤¼ĒĢ£ ņ£ĀņĀäņĀü ņåīņØĖņØä ĒåĄĒĢ£ ņØ┤ļÅģņä▒ ļé£ņ▓Ł ļ░£ņāØ ņśłņĖĪļ╣äņ£©ņØ┤ ņāüļīĆņĀüņ£╝ļĪ£ ļåÆņ¦Ć ņĢŖņ£╝ļ®░, ņśżĒ׳ļĀż ņל ņĢīļĀżņĀĖ ņ׳ļŖö ņØ┤ļÅģņä▒ ņĢĮņĀ£ņØĖ ņĢäļ»ĖļģĖĻĖĆļ”¼ņĮöņé¼ņØ┤ļō£ņÖĆ ņØ┤ļć©ņĀ£ ļō▒ņØś ņĢĮņĀ£ Ēł¼ņŚ¼ļéś ļæÉĻ▓ĮļČĆ ļ░®ņé¼ņäĀ ņĪ░ņé¼ļź╝ ļÅÖļ░śĒĢ£ Ļ▓ĮņÜ░ ņåīņØī ļģĖņČ£ ļō▒ņŚÉņä£ ņØ┤ļÅģņä▒ ļé£ņ▓Ł ļ╣äņ£©ņØ┤ ļåÆņØīņØ┤ ņĢīļĀżņĀĖ ņ׳ļŗż[59,93,94].
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØä ļ¦ēĻĖ░ ņ£äĒĢ£ ņŚ░ĻĄ¼
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØä ņśłļ░®ĒĢśĻĖ░ ņ£äĒĢ┤ņä£ļŖö Ļ│ĀņŻ╝Ēīī ņ▓ŁļĀźĻ▓Ćņé¼ ļ░Å distortion product otoacoustic emission (DPOAE) ļō▒ņØś ņĀĢĻĖ░ņĀüņØĖ Ļ▓Ćņé¼ļź╝ ĒåĄĒĢ£ ļé£ņ▓ŁņØś ņĪ░ĻĖ░ ļ░£Ļ▓¼ņØ┤ ļÅäņøĆņØ┤ ļÉśļ®░, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņØä Ļ░Éņåīņŗ£ĒéżļŖö ļ░®ļ▓Ģņ£╝ļĪ£ Ļ│ĀņןņĢĪ ņŗØņŚ╝ņłśņØś ņĀäņ▓śņ╣śĻ░Ć ĒĢäņÜöĒĢ£ Ļ▓ĮņÜ░ ļÅģņä▒ņØ┤ ņĀüņØĖ 2ņäĖļīĆ ņĢĮļ¼╝ņØĖ carboplatinņ£╝ļĪ£ņØś ļīĆņ▓┤ Ēł¼ņŚ¼ļź╝ ĒĢśĻĖ░ļÅä ĒĢ£ļŗż. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ĒĢŁņĢöņĀ£ņØś ņØ┤ļÅģņä▒ ĻĖ░ņĀäņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļōżņØ┤ ņ¦äĒ¢ēļÉśļ®┤ņä£, ņäĖĒż ņé¼ļ®ĖņØś ņ¢ĄņĀ£, ĒÖ£ņä▒ĒÖö ņé░ĒśĖņØś ņżæĒÖö, ņŚ╝ņ”Ø ļ░śņØæņØś ņ¢ĄņĀ£ ļō▒ ņ×Āņ×¼ņĀü ļ│┤ĒśĖ Ļ░ĆļŖźņä▒ņØä ņ¦Ćļŗī ņĢĮļ¼╝ņØś ņŗ£ļÅäņÖĆ ņŚ░ĻĄ¼Ļ░Ć ņ׳ņŚłļŗż. ļīĆļČĆļČäņØś ņĀäņ×äņāü ļ░Å ņ×äņāüņŗ£ĒŚśņŚÉņä£ņØś ņØ┤ļÅģņä▒ ļ│┤ĒśĖ ņĢĮļ¼╝ņØĆ ĒĢŁņé░ĒÖöņĀ£ ļ░Å ĒĢŁņŚ╝ņ”ØņĀ£ ļ▓öņŻ╝ņŚÉ ņåŹĒĢśļ®░, ņØ┤ņŚÉ Ļ┤ĆļĀ©ĒĢ£ Ļ▓ĮļĪ£ ņżæ ĒĢśļéś ļśÉļŖö ļæś ļ¬©ļæÉļź╝ ņ¢ĄņĀ£ĒĢśļŖö ĻĖ░ņĀäņ£╝ļĪ£ ņ×æņÜ®ĒĢ£ļŗż. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØä ņŻ╝ņ×ģĒĢ£ ņŚ¼ļ¤¼ ļÅÖļ¼╝ ļ¬©ļŹĖņŚÉņä£ ĒĢŁņé░ĒÖöņĀ£ņØĖ apocynin, lactage, alpha lipoic acids ļō▒, ĒĢŁņŚ╝ņ”Ø ņ×æņÜ®ņØä ļÅÖņŗ£ņŚÉ ĒĢśļŖö flunarizine, hydrogen, N-acetylcystein ļ░Å ņäĖĒżņ×Éļ®Ėņé¼ņŚÉ ņśüĒ¢źņØä ņŻ╝ļŖö pifithrin-a, minocycline ļō▒ņØś ņ▓śļ”¼ņŗ£ ņ▓ŁļĀź ņŚŁņ╣śņØś ņāüņŖ╣ ļ░Å ņ£Āļ¬©ņäĖĒż ņåÉņāüņØś Ļ░Éņåīļź╝ ĒÖĢņØĖĒĢśņśĆļŗż(Table 2). ņ×äņāüņŚÉņä£ļŖö thiamin, cyanin ļ░Å ņŗżĒŚśņĀüņ£╝ļĪ£ sodium thiosulfate, lazaroid ļō▒ņØ┤ ņé¼ņÜ®ļÉśĻ│Ā ņ׳ņ£╝ļéś, Ēśäņ×¼Ļ╣īņ¦Ć FDA ņŖ╣ņØĖņØä ļ░øņØĆ ņĢĮļ¼╝ņØĆ ņŚåļŖö ņāüĒā£ņØ┤ļŗż.
Ļ▓░ ļĪĀ
ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØĆ ņĢöĒÖśņ×ÉņØś ņāØļ¬ģņØä ĻĄ¼ĒĢśļŖö ļīĆĒæ£ņĀüņØĖ ĒĢŁņĢöņĀ£ļĪ£ņä£ņØś ņŚŁĒĢĀņØä ĒĢśņ¦Ćļ¦ī ļ╣äĻ░ĆņŚŁņĀü Ļ░ÉĻ░üņŗĀĻ▓Įņä▒ ļé£ņ▓ŁņØä ņ£Āļ░£ĒĢśļŖö ņØ┤ļÅģņä▒ ļśÉĒĢ£ ļé┤ĒżĒĢśļŖö ņĢĮņĀ£ņØ┤ļŗż. ņŚ╝ņ”ØņØĆ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ļģĖņČ£ Ēøä ņ▓śņØīņ£╝ļĪ£ ņ┤ēļ░£ļÉśļŖö ļ░śņØæņØ┤ļ®░, ņé░ĒÖöņŖżĒŖĖļĀłņŖżņÖĆ ROS ņāØņä▒ņ£╝ļĪ£ ņØ┤ņ¢┤ņ¦ĆļŖö ņØ╝ļĀ©ņØś ļ░śņØæņØä ņ£Āļ░£ĒĢ£ļŗż. ROSļŖö ņ¦Ćņ¦łņØś Ļ│╝ņé░ĒÖö, ļŗ©ļ░▒ņ¦ł ņ¦łĒÖö, DNA ļ│ĆĻ▓Į ļ░Å ņŚ╝ņ”Ø Ļ│╝ņĀĢņØś ņ”ØĒÅŁņØä ĒåĄĒĢ┤ ņäĖĒż ņåÉņāüņØä ņØ╝ņ£╝Ēé¼ ņłś ņ׳ņ£╝ļ®░, ņÖĆņÜ░ņäĖĒżļŖö ļŗ©ļ░▒ņ¦ł ļ░Å DNA ļ│ĄĻĄ¼ ļ®öņ╗żļŗłņ”śņØä ĒÖ£ņä▒ĒÖöĒĢśņŚ¼, ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ļÅģņä▒ņ£╝ļĪ£ļČĆĒä░ ĒĢŁņāüņä▒ņØä ĒÜīļ│ĄĒĢśĻĖ░ ņ£äĒĢ£ ļģĖļĀźņØä ĒĢ£ļŗż. ĻĘĖļ¤¼ļéś, ROSļĪ£ ņØĖĒĢ┤ ņ£ĀļÅäļÉ£ ņäĖĒżņØś ņåÉņāüņŚÉ ļīĆĒĢ£ ļ░®ņ¢┤ ņŗ£ņŖżĒģ£ņØś ņŗżĒī©ļŖö Ļ▓░ĻĄŁ ņäĖĒż ņé¼ļ®ĖļĪ£ ņØ┤ņ¢┤ņ¦ĆĻ▓ī ļÉśļ®░, ļé┤ņØĖņä▒ ņäĖĒżņ×Éļ®Ėņé¼, pyroptosis, ņ×ÉĻ░ĆĒżņŗØ ļ░Å necroptosis ļśÉĒĢ£ ņØ┤ņŚÉ Ļ┤ĆņŚ¼ĒĢĀ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
Ēśäņ×¼Ļ╣īņ¦Ć ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ņŚÉ ļīĆĒĢ£ ļŗżņ¢æĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņ¦äĒ¢ēļÉśņŚłņ£╝ļéś, ņŚ░ĻĄ¼ Ļ▓░Ļ│╝ņØś ļģ╝ļ×ĆņØ┤ ņ׳Ļ▒░ļéś ļČłņÖäņĀäĒĢ£ Ļ▓░Ļ│╝ļź╝ ļ│┤Ļ│ĀĒĢśņśĆĻ│Ā, ņØ┤ļÅģņä▒ņŚÉ ļīĆĒĢ£ ļ│┤ĒÄĖņĀüņØĖ ņ▓ŁļĀź ĒÅēĻ░Ć ļ░Å ņØ┤ļÅģņä▒ ļō▒ĻĖēņŚÉ ļīĆĒĢ£ Ēæ£ņżĆĒÖöļÉ£ ņ¦Ćņ╣©ņØ┤ ņŚåļŖö ņāüĒā£ļĪ£, ņ×äņāü ņŚ░ĻĄ¼ņØś Ļ▓░Ļ│╝ņØś ĒĢ┤ņäØņŚÉ ņ׳ņ¢┤ņä£ļÅä ļģ╝ļ×ĆņØ┤ ņ׳ļŖö ņāüĒā£ņØ┤ļŗż.
ļö░ļØ╝ņä£ Ē¢źĒøä ņŚ░ĻĄ¼ļŖö ņŗ£ņŖżĒöīļØ╝Ēŗ┤ņØś ņ╣śļŻī ĒÜ©ļŖźņØä ļ░®ĒĢ┤ĒĢśņ¦Ć ņĢŖņ£╝ļ®┤ņä£, ļŗżņ¢æĒĢ£ ĒÖ£ņä▒ Ļ▓ĮļĪ£ļź╝ ņ¢ĄņĀ£ĒĢśļŖö ļ│ĄĒĢ® ņĢĮļ¼╝ ņé¼ņÜ® ļ░Å ĒÜ©Ļ│╝ņĀüņØĖ ĻĄŁņåī ņÜöļ▓ĢņØś Ļ░£ļ░£ņØä Ļ│ĀļĀżĒĢśļŖö Ļ▓āņØ┤ ĒĢäņÜöĒĢĀ ņłś ņ׳ļŗż. ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņØ┤ļÅģņä▒ ĻĖ░ņĀäņŚÉ ļīĆĒĢ£ ļŹö Ļ╣ŖņØĆ ņØ┤ĒĢ┤ļŖö Ē¢źĒøä ņŚ░ĻĄ¼ņ×ÉļōżņØ┤ ņŗ£ņŖżĒöīļØ╝Ēŗ┤ ņĢĮļ¼╝ ņ×æņÜ® ĻĖ░ņĀäņØä ļ╣äĒÖ£ņä▒ĒÖöĒĢśļŖö Ēæ£ņĀü ņŚ░ĻĄ¼ļź╝ ĒĢśļŖö ļŹ░ ļÅäņøĆņØ┤ ļÉĀ ņłś ņ׳ņØä Ļ▓āņØ┤ļŗż.