Benzyl Isothiocyanate Suppresses Myofibroblast Differentiation and Extracellular Matrix Production in TGF-β1–Induced Nasal Polyp Fibroblasts
TGF-β1 유도 비용종 섬유모세포에서 Benzyl Isothiocyanate의 근섬유모세포의 분화 및 세포외 기질 생산의 억제 작용에 대한 연구
Article information

Abstract
Background and Objectives
Benzyl isothiocyanate (BITC) is an effector molecule in many cruciferous vegetable defense systems with antioxidant, anti-tumor, and anti-inflammatory activity. In the present study, the effects of BITC on organ-cultured nasal polyps (NPs) and transforming growth factor (TGF)-β1-induced profibrotic responses in nasal polyp-derived fibroblasts (NPDFs) were evaluated, and the associated signaling pathways were explored.
Materials and Method
Organ-cultured NPs and NPDFs were obtained from patients with NPs. Organ-cultured NPs were treated with BITC (0 or 5 μM). The NPDFs were treated with TGF-β1 (1 ng/mL) in the presence or absence of BITC (1, 3, and 5 μM). The expression levels of Collagen-1 (Col-1) and α-smooth muscle actin (α-SMA) in organ-cultured NPs were measured by western blot analysis. The expression levels of Col-1, α-SMA, and phosphorylatedsmall mothers against decapentaplegic (Smad2 and Smad3) in NPDFs were also measured by western blot analysis. The contractile activity of the NPDFs was determined by a collagen gel contraction assay.
Results
BITC reduced α-SMA and Col-1 expression in organ-cultured NPs but did not show biologically meaningful cytotoxic effects in TGF-β1-stimulated NPDFs within the tested concentration range. In TGF-β1-stimulated NPDFs, BITC attenuated the expression of α-SMA and Col-1 and suppressed collagen gel contraction. BITC treatment was associated with decreased phosphorylation of Smad2 and Smad3.
Conclusion
These findings suggest that BITC may exert antifibrotic effects in TGF-β1-induced fibroblast activation and may serve as a potential modulator in fibrosis-dominant remodeling of NPs.
Introduction
Nasal polyps (NPs) are characterized by the accumulation of inflammatory cells, thickening of the basement membrane, abnormal proliferation of fibroblasts, and excessive deposition of the extracellular matrix [1]. Fibroblasts contribute to the structural formation and development of NPs by providing extracellular matrix proteins such as collagen, fibronectin, and vi mentin. In addition to this profibrotic role, fibroblasts are suggested to possess inflammatory functions that may play important roles in the pathogenesis and persistence of NPs [2].
Myofibroblasts are defined by their morphological features, functional properties, and gene expression patterns. They serve as key effector cells that synthesize large amounts of extracellular matrix proteins, including collagen-1 (Col-1) and fibronectin, along with profibrotic proteins, particularly α-smooth muscle actin (α-SMA). Myofibroblasts play important roles in extracellular matrix remodeling across various pathological conditions of the human airway, including asthma, chronic rhinosinusitis, and NPs [3].
Although multiple factors are involved in the development and progression of fibrosis, transforming growth factor-β (TGF-β) is considered one of the most potent fibrogenic mediators and a key regulator in the pathophysiology of fibrosis. TGF-β is a secreted homodimeric protein that regulates diverse biological processes, including cell proliferation, differentiation, migration, extracellular matrix production, angiogenesis, and apoptosis [4,5]. The overexpression of TGF-β has been associated with fibrotic diseases in various human organs, including pulmonary fibrosis, cardiac fibrosis, liver cirrhosis, glomerulosclerosis, diabetic nephropathy, Crohn’s disease, rheumatoid arthritis, radiation-induced fibrosis, and myocarditis [6]. TGF-β1 is well known to activate fibrotic responses mainly through the small mothers against decapentaplegic (Smad) signaling pathway. Smad proteins are essential components of the TGF-β family signaling pathway, and the regulation of this pathway may represent an effective therapeutic strategy for NPs. Furthermore, TGF-β1 plays an important role in the pathophysiology of NPs by inducing myofibroblast differentiation and increasing extracellular matrix accumulation [7].
Organ culture preserves the structural integrity of cells and stroma in an organ-specific manner while maintaining tissue architecture, making it a more clinically relevant ex vivo model [8]. Isothiocyanates are natural compounds abundant in cruciferous vegetables such as broccoli, watercress, and Brussels sprouts. Specific isothiocyanates, including sulforaphane, phenethyl isothiocyanate (PEITC), and benzyl isothiocyanate (BITC), are derived from glucosinolates present in these vegetables [9]. BITC exhibits antioxidant, anti-tumor, and anti-inflammatory activities, functioning as an effector molecule in the defense mechanisms of various cruciferous plants [10-12].
However, there have been no prior reports indicating that BITC exerts antifibrotic effects during NP formation. In the present study, we investigated the effects of BITC on profibrotic responses and their related signaling pathways in both organ-cultured NPs and TGF-β1-induced nasal polyp-derived fibroblasts (NPDFs).
Materials and Methods
Patients with NPs were recruited from the Department of Otorhinolaryngology at Inje University Busan Paik Hospital. Written informed consent for participation was obtained from all patients, and the study protocol was approved by the Institutional Review Board (IRB approval no. 2020-0034).
NPs were defined as polyps growing into the nasal cavity from both middle meatuses upon endoscopic examination, with the involvement of the ethmoid and maxillary sinuses confirmed via sinus computed tomography. All polyps were resected from the middle meatus at the beginning of surgery. Patients who had received systemic or topical steroids, nonsteroidal anti-inflammatory drugs, antihistamines, or macrolide antibiotics within 4 weeks prior to the biopsy were excluded. None of the subjects had a history of rhinitis, asthma, or aspirin hypersensitivity. Allergy diagnoses were based on patient history combined with the results of ImmunoCAP or skin prick testing.
Reagents
BITC and the selective inhibitor of Smad3 (SIS3) were purchased from Sigma-Aldrich (St. Louis). TGF-β1 was obtained from R&D Systems, and the Cell Counting Kit-8 (CCK-8) was purchased from Dojindo Laboratories. Antibodies against α-SMA (cat. no. ab5694) and Col-1 (cat. no. ab88147) were supplied by Abcam Inc., while the antibody against actin (cat. no. 612656) was acquired from BD Biosciences. The antibody against glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (cat. no. LF-PA0018) was purchased from YoungIn Frontier. Antibodies against Smad2 (cat. no. 5339), phosphorylated Smad2 (p-Smad2) (cat. no. 3101), Smad3 (cat. no. 9523), and phosphorylated Smad3 (p-Smad3) (cat. no. 9520) were procured from Cell Signaling Technology Inc.
Organ culture of NPs
A total of 6 NP tissues (male to female ratio=2:4, median age 41 years) were cut into 2 to 3 mm3 pieces under sterile conditions using a surgical blade. The tissue pieces were washed three times with phosphate-buffered saline (PBS) containing fungizone (5 μM) and penicillin G (300 μM). They were then rinsed with Dulbecco’s Modified Eagle Medium (DMEM) supplemented with calf serum (10%) and gentamicin (20 μM). The tissue fragments were placed on hydrated gelatin sponges (1×1 cm, Spongostan; Johnson & Johnson) with the mucosal side facing upward and the submucosal side facing downward. Each sponge was placed into a well of a 6-well plate, and 3 mL of culture medium was added to ensure the mucosal surface remained above the liquid line. The plates were cultured at 37°C in 5% CO2. BITC (0 or 5 μM) was added to the culture medium according to the experimental conditions. The tissues were divided into two groups: a control group preincubated in regular culture medium for 1 hour and then organ-cultured for 24 hours, and a 5 μM BITC-treated group preincubated in culture medium containing 5 μM BITC for 1 hour and then organ-cultured for 24 hours in a medium containing the same concentration of BITC.
Culture of NPDFs
NPDFs were isolated from the surgically obtained tissues through enzymatic digestion using collagenase (500 U/mL; Sigma-Aldrich, Merck KGaA), hyaluronidase (30 U/mL; Sigma-Aldrich, Merck KGaA), and DNase (10 U/mL; Sigma-Aldrich, Merck KGaA). The isolated cells were cultured in DMEM containing 10% (v/v) heat-inactivated fetal bovine serum (Invitrogen, Thermo Fisher Scientific Inc.), penicillin (1000 U/mL), and streptomycin (1000 μM; Invitrogen, Thermo Fisher Scientific Inc.) at 37°C in 5% CO2. The purity of the NPDFs was confirmed via flow cytometry and observation of the spindle-shaped morphology characteristic of fibroblasts. The cells were utilized for experiments after 4 to 6 passages.
Measurement of cell viability
Cell viability was assessed using the CCK-8 reagent. NPDFs (1×105 cells/well) were seeded into 96-well microplates and treated with BITC at concentrations of 5, 10, and 30 μM. Following a 24-hour incubation at 37°C in 5% CO2, the cells were washed twice with PBS. The CCK-8 solution was added to each well, and the cells were incubated at 37°C for 1 hour. Absorbance was measured at 450 nm using a microplate reader (SpectraMax M2e; Molecular Devices LLC). All experiments were performed in triplicate.
Western blot analysis
Cells were lysed using Mammalian Cell-PE LB lysis buffer (G-Biosciences). Protein concentrations were quantified using the Bradford assay (Bio-Rad protein assay dye reagent; Bio-Rad Laboratories Inc.). A total of 20 μg of protein was separated on 10% SDS-polyacrylamide mini gels and transferred to nitrocellulose membranes (GE Healthcare Life Sciences). The membranes were blocked for 1 hour at room temperature using a blocking solution containing 5% non-fat milk in Tris-buffered saline with 0.1% Tween-20 (TBST). The membranes were then incubated overnight at 4°C with primary antibodies against α-SMA, Col-1, p-Smad2, and pSmad3, each diluted at 1:1000. Following this, they were incubated for 1 hour at room temperature with a horseradish peroxidase-conjugated secondary antibody (goat anti-mouse IgG, diluted 1:1000). After washing three times with TBST, the immunoreactive bands were visualized using an electrochemiluminescence immunoassay (Pierce; Thermo Fisher Scientific Inc.). Band intensities were quantified using Multi Gauge software (version 2.2; Fuji Film)
Rat Tail Type-I Collagen Gel Contraction Assay
Rat tail type-I collagen was diluted with fibroblast basal medium (CC-3131; Lonza Group Ltd.) to a final concentration of 1 mg/mL and mixed with NPDFs to achieve a final cell concentration of 1×105 cells/mL. Following the addition of 1 N NaOH according to the manufacturer’s protocol, 500 μL of the cell-collagen mixture was dispensed into each well of a 24-well cell culture plate. The plate was incubated at 37°C for 30 minutes to facilitate gel formation, after which the medium was replaced with fibroblast growth medium 2 and cultured overnight. BITC and TGF-β1 were then applied based on the experimental conditions. Gel sizes were measured using ImageJ software (version 1.51j8; National Institutes of Health).
Statistical analysis
All data are presented as the mean±standard error of the mean (SEM). Statistical analyses were performed using GraphPad Prism software (version 5.0; GraphPad Software Inc.). Comparisons between groups were conducted using Dunnett’s multiple range test. A p-value of <0.05 was considered to indicate statistical significance.
Results
Effects of BITC on α-SMA and Col-1 production in organ-cultured NPs
To determine whether BITC suppresses α-SMA and Col-1 protein expression in NPs, organ cultures were performed using collected NP tissues. The results demonstrated that BITC significantly inhibited the production of both α-SMA and Col-1 proteins in the organ-cultured NPs (Fig. 1).
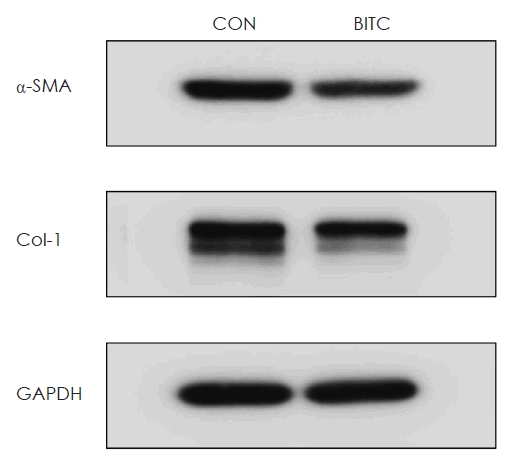
Effect of BITC on α-SMA and Col-1 protein expression in nasal polyp organ cultures. Nasal polyp tissues were cultured and stimulated with (5 μM) or without BITC. The protein expression level of α-SMA, Col-1, and GAPDH were examined using western blotting. Expression of a housekeeping gene, GAPDH was used as an internal control. BITC, benzyl isothiocyanate; α-SMA, α-smooth muscle actin; Col-1, collagen-1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Effect of BITC on the viability of NPDFs
The viability of BITC-treated NPDFs was evaluated using the CCK-8 assay. Overall, cell viability remained high across the BITC concentration range of 1 to 5 μM, although statistically significant decreases were observed at the 3 μM and 5 μM levels (Fig. 2). However, viability at these concentrations remained above 80% to 90%, which did not indicate biologically meaningful cytotoxicity at these doses. Conversely, viability decreased markedly at concentrations of 10 μM or higher. Based on these findings, the BITC concentration range for all subsequent functional experiments was established at 1 to 5 μM.
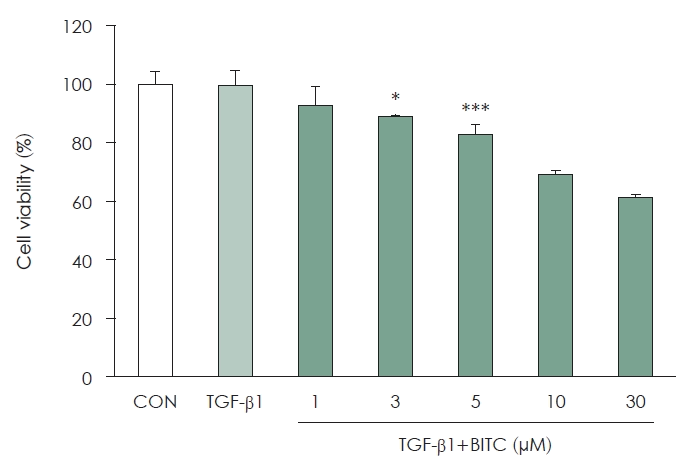
Effects of BITC on the viability of TGF-β1–stimulated NPDFs. NPDFs were pretreated with increasing concentrations of BITC (1, 3, 5, 10, and 30 μM) for 30 min and subsequently stimulated with TGF-β1 (1 ng/mL). Cell viability was assessed using the Cell Counting Kit-8 assay. Results are expressed as percentages relative to the TGF-β1–stimulated group. Data are presented as the mean±SEM from three independent experiments. *p<0.05 and ***p<0.001 vs. TGF-β1–stimulated group. BITC, benzyl isothiocyanate; TGF, transforming growth factor; NPDF, nasal polyp–derived fibroblasts.
Effects of BITC on α-SMA and Col-1 production in TGF-β1-stimulated NPDFs
To determine whether BITC suppresses Col-1 and α-SMA expression in TGF-β1-stimulated NPDFs, the cells were pretreated with BITC (1 to 5 μM) for 30 minutes prior to being treated with TGF-β1 for 24 hours. The expression levels of Col-1 and α-SMA decreased significantly in a BITC dosedependent manner (Fig. 3).
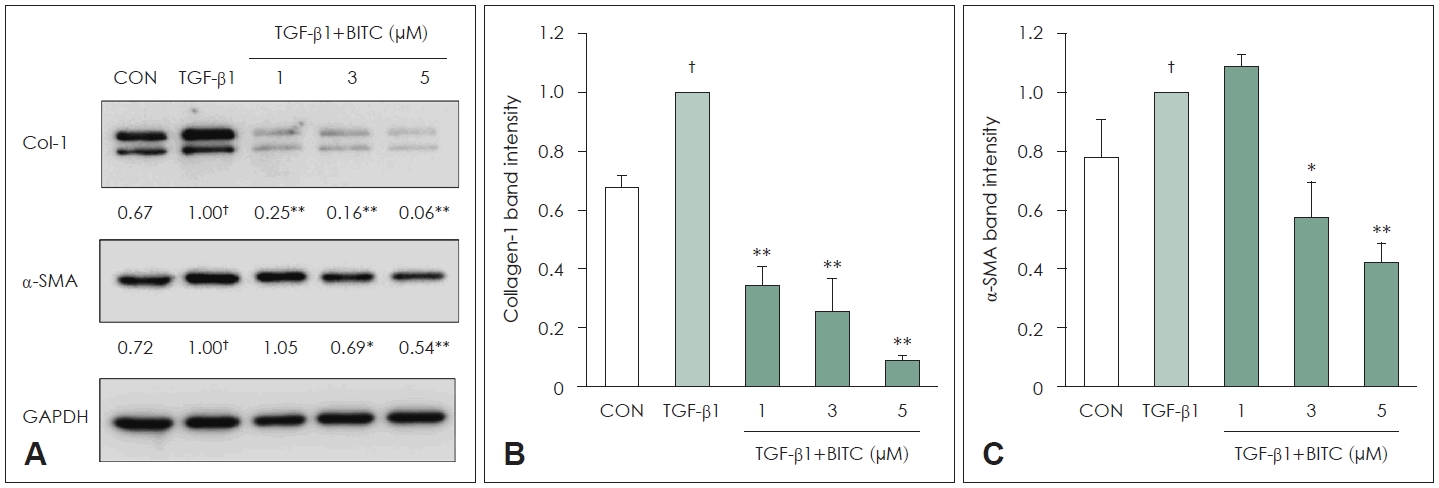
Effects of BITC on α-SMA and Col-1 protein expression in TGF-β1-stimulated NPDFs. A: Representative western blot images showing α-SMA and Col-1 protein expression following BITC pretreatment (1, 3, and 5 μM) and subsequent TGF-β1 stimulation (1 ng/mL). GAPDH was used as a loading control. B and C: Densitometric analyses of Col-1 (B) and α-SMA (C) protein expression. Band intensities were normalized to GAPDH and expressed relative to the TGF-β1–stimulated group. Each value indicates the mean±SEM and is representative of results obtained from three independent experiments. †p<0.05 vs. control group (CON, no treatment); *p<0.05 and **p<0.01 vs. TGF-β1-stimulated group. BITC, benzyl isothiocyanate; α-SMA, α-smooth muscle actin; TGF, transforming growth factor; Col-1, collagen-1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NPDF, nasal polyp–derived fibroblasts.
BITC inhibits the TGF-β1-induced Smad2 and Smad3 signaling pathways
In NPDFs, stimulation with TGF-β1 markedly increased the phosphorylation of Smad2 and Smad3. However, when the cells were pretreated with BITC (1 to 5 μM) for 30 minutes prior to a 24-hour TGF-β1 treatment, the phosphorylation levels of Smad2 and Smad3 were significantly reduced. Thus, the antifibrotic effects of BITC appear to be mediated, at least in part, by inhibition of TGF-β1-induced Smad2/3 signaling (Fig. 4). Furthermore, blocking the Smad pathway using SIS3 effectively reduced the expression of Col-1 and α-SMA proteins (Fig. 5).
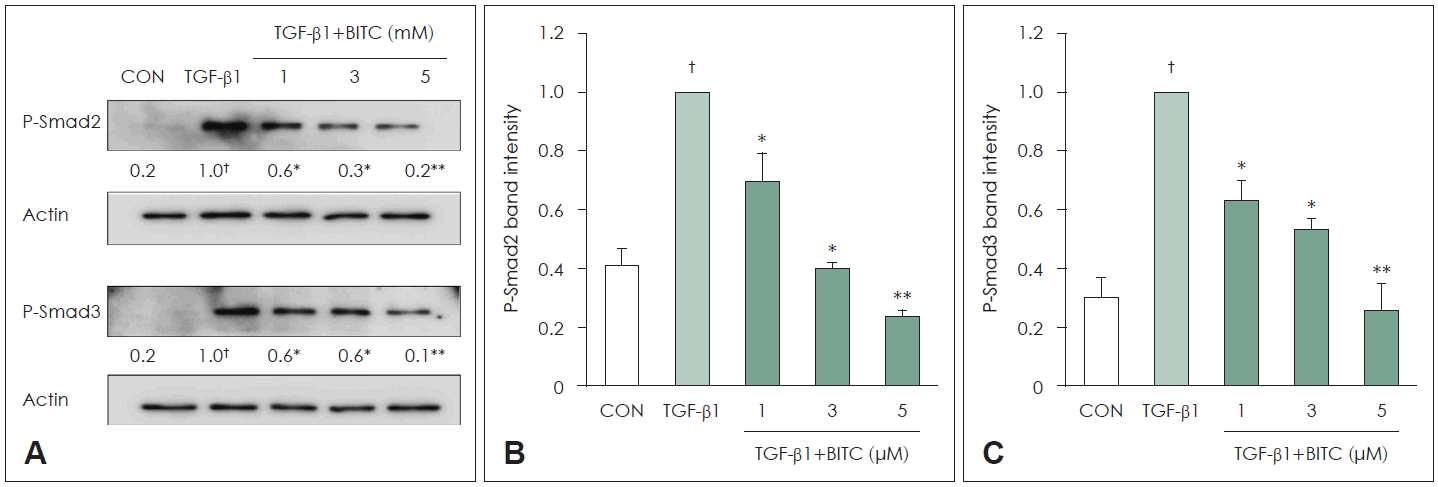
Effects of BITC on Smad2 and Smad3 activation in TGF-β1–stimulated NPDFs. A: Representative western blot images showing phosphorylation levels of Smad2 (p-Smad2) and Smad3 (p-Smad3) following BITC pretreatment (1, 3, and 5 μM) and subsequent TGF-β1 stimulation (1 ng/mL). Actin was used as a loading control. B and C: Densitometric analyses of p-Smad2 (B) and p-Smad3 (C) protein expression. Band intensities were normalized to Actin and expressed relative to the TGF-β1–stimulated group. NPDFs were pretreated with BITC for 30 min and then stimulated with TGF-β1 for 30 min. Each value indicates the mean±SEM and is representative of results obtained from three independent experiments. †p<0.05 vs. control group (CON, no treatment); *p<0.05 and **p<0.01 vs. TGF-β1-stimulated group. BITC, benzyl isothiocyanate; TGF, transforming growth factor; NPDF, nasal polyp–derived fibroblasts.
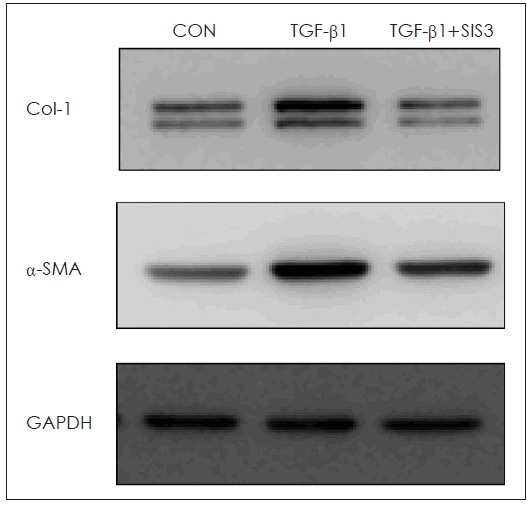
Effect of Smad on the expression of α-SMA and Col-1 proteins in TGF-β1-stimulated NPDFs. The cells were seeded at 2×105 cells/mL and incubated with SIS3 (Smad 3 inhibitor) for 1 hour prior to TGF-β1 stimulation (1 ng/mL). Following stimulation with TGF-β1 for 3 hours, the α-SMA protein and Col-1 protein expression was determined via western blotting. GAPDH was used as an internal control. α-SMA, α-smooth muscle actin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; TGF, transforming growth factor; Col-1, collagen-1; NPDF, nasal polyp–derived fibroblasts; SIS3, selective inhibitor of Smad3.
Effect of BITC on fibroblast contractile activity
A type I collagen gel contraction assay was conducted to evaluate whether BITC blocks the TGF-β1-induced contractile activity of NPDFs. Following TGF-β1 stimulation, the collagen gel size was significantly reduced to 82.44% relative to the group not treated with TGF-β1 (100%). However, pretreatment with BITC effectively inhibited this contractile effect, resulting in gel sizes of 86.35%, 103.74%, and 118.09% at BITC concentrations of 1, 3, and 5 μM, respectively (Fig. 6).
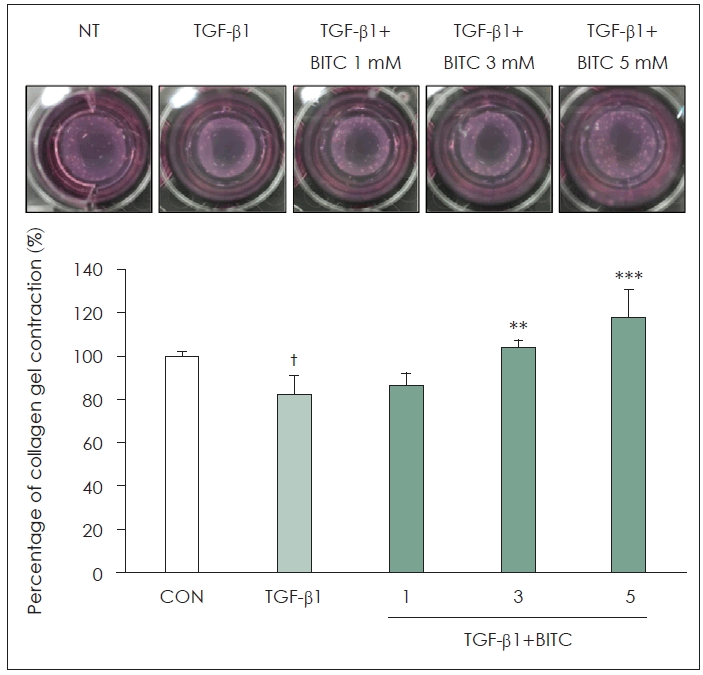
Effects of BITC on the contractile activity of NPDFs. TGF-β1 induction decreased the size of the gel, while BITC pretreatment inhibited this reduction. Data is presented in the form of the percentage of gel surface area in each well to the control gel surface area. †p<0.05 vs. control group (CON, no treatment); **p<0.01 and ***p<0.001 vs. TGF-β1-stimulated group. BITC, benzyl isothiocyanate; NPDF, nasal polyp–derived fibroblasts; TGF, transforming growth factor.
Discussion
The formation of NPs has been proposed to initiate with an injury to the mucosal epithelium, which is followed by the accumulation of extracellular matrix proteins and matrix metalloproteinases, as well as the infiltration of various inflammatory cells [13]. BITC has been reported to inhibit cell proliferation and to exert both anticancer and anti-inflammatory activities [12,14,15]. Its anticancer effects are believed to occur primarily through the suppression of proteins involved in tumor initiation and proliferation, thereby inhibiting the overall growth of cancer cellsn [15].
Microbial biofilms have additionally been implicated in the pathophysiology of NPs.16) Biofilms are defined as microbial communities embedded within extracellular polymeric substances that are irreversibly attached to abiotic or biotic surfaces. Bacteria organized into these multilayered structures differ both phenotypically and genotypically from their planktonic counterparts. Moreover, biofilms provide an ideal environment for the active exchange of genes involved in antimicrobial resistance [17,18]. BITC has been documented to significantly inhibit biofilm formation by Staphylococcus aureus [19].
Interleukin (IL)-13 is a representative Th2-derived cytokine that acts as an important mediator in chronic inflammation and tissue remodeling [20]. BITC may exert anti-inflammatory and antifibrotic effects not only through its antimicrobial actions but also via distinct immunologic pathways. IL-13 is known to induce goblet cell hyperplasia, inducible nitric oxide synthase production, and fibrosis, thereby amplifying inflammation and promoting tissue remodeling [20,21]. In NP tissues, IL-13 mRNA expression was found to be significantly higher than in controls, and this overexpression is likely as-sociated with NP development [22]. Notably, BITC significantly suppressed IL-13 expression in human basophilic KU812 cells stimulated with calcium ionophore [23]. Beyond these antiinflammatory and antimicrobial properties, the present study focused on whether BITC also modulates fibrotic remodeling in NP tissues.
Given that no established in vivo model for NPs is currently available, we utilized an organ culture model based on the air-liquid interface technique. This model successfully preserves both submucosal and epithelial tissue components while maintaining structural integrity and biochemical activity. It also sustains the viability of the major cellular components of NPs, particularly eosinophils, which are highly vulnerable to injury. Additionally, the ability of various cells, including surface epithelial cells, inflammatory cells, and endothelial cells, to express vascular endothelial growth factor (which is involved in NP formation) is well preserved in this model [24]. In the present study, BITC significantly inhibited the production of α-SMA and Col-1 proteins in the organ-cultured NP tissues.
Fibroblasts act as central mediators of extracellular matrix accumulation, cellular differentiation, and proliferation in response to persistent tissue injury [25]. Such persistent tissue injury frequently induces the differentiation of fibroblasts into myofibroblasts, which subsequently participate in the broader inflammatory response to the injury [26].
It is well established that TGF-β induces fibroblast activation, proliferation, and differentiation. Previous studies investigating NPDFs have demonstrated that exposure to TGF-β1 induces differentiation into myofibroblasts, increases collagen production, and enhances α-SMA expression [27,28]. Consistent with these findings, TGF-β1 stimulation significantly increased the expression of α-SMA and Col-1 in NPDFs in the current study. However, treatment with BITC at concentrations ranging from 1 to 5 μM significantly and dose-dependently suppressed the TGF-β1-induced increases in α-SMA and Col-1 expression without causing cytotoxicity. These findings suggest that BITC attenuates the TGF-β1-induced increase in α-SMA and Col-1 production by actively inhibiting TGF-β1-related signaling pathways. Smad-dependent transcription is essential for TGF-β signaling, and this pathway represents one of the most common mechanisms operating during fibrosis. When TGF-β1 binds to its receptor, Smad2 or Smad3 becomes phosphorylated. It then binds to Smad4 and translocates into the nucleus. These resultant complexes activate the transcription of fibrosis-related genes within the nucleus, thereby inducing fibrotic changes [29].
In the present study, BITC treatment dose-dependently reduced the TGF-β1-induced phosphorylation of Smad2 and Smad3, indicating that the antifibrotic effects of BITC are linked to the Smad signaling pathway. Furthermore, blocking the Smad pathway using a specific inhibitor such as SIS3 effectively reduced the expression of Col-1 and α-SMA proteins. Finally, we evaluated the effects of BITC on the type I collagen gel contraction induced in TGF-β1-stimulated NPDFs. A model in which fibroblasts are cultured within a three-dimensional collagen gel has been widely utilized as a tissue contraction model that effectively mimics the in vivo environment and characterizes active fibrosis [30]. Fibroblasts pretreated with BITC exhibited reduced contractile activity compared with cells treated with TGF-β1 alone, as confirmed by the significant attenuation of collagen gel contraction following BITC treatment. These results suggest that BITC may inhibit the responses associated with myofibroblast differentiation and extracellular matrix production in TGF-β1-stimulated NPDFs.
This study has several limitations. First, the number of NP tissues utilized for the organ cultures was limited (n=3 to 6), which may not adequately reflect the high interindividual variability inherently present in human-derived samples. Consequently, statistical interpretation may be constrained, and the current findings should be interpreted with appropriate caution. Second, no histologic or molecular analyses were included to distinguish the specific inflammatory endotypes of the NPs, and no stratified analyses were performed according to clinical characteristics such as eosinophil infiltration levels or the presence of concurrent asthma. The pathogenesis, tissue responses, and treatment responses of NPs can differ significantly according to the underlying inflammatory endotype and clinical background. Therefore, these findings may not be generalizable to all patients presenting with chronic rhinosinusitis with NPs. Third, no genetic manipulation or selective inhibition experiments were performed to robustly distinguish the individual roles of Smad2 and Smad3. Furthermore, no analyses were included to directly evaluate the nuclear translocation of these Smad proteins. Further comprehensive studies are therefore required to more precisely clarify the antifibrotic effects of BITC and to validate the related mechanisms proposed in this study.
In this study, BITC effectively inhibited the expression of fibrotic markers such as α-SMA and Col-1, as well as collagen gel contraction, in vitro in TGF-β1-stimulated NPDFs. These inhibitory effects may be associated with the suppression of the Smad2 and Smad3 signaling pathways. Given that multiple pathophysiologic mechanisms are involved in NP formation and tissue remodeling, and that fibrotic responses represent merely one component of this complex process, the present findings suggest that BITC may act as a potential therapeutic modulator in specific, limited contexts, such as in NP subtypes with relatively predominant fibrotic responses or within settings of postoperative recurrence.
Supplementary Materials
Korean translation of this article is available with the Online-only Data Supplement at https://doi.org/10.3342/kjorl-hns.2025.00605.
Notes
Acknowledgments
None
Author Contribution
Conceptualization: Seong Kook Park. Data curation: Sung Yool Park. Formal analysis: Sung Yool Park. Investigation: Seong Kook Park. Methodology: Sung Yool Park. Project administration: Sung Yool Park. Resources: Sung Yool Park. Software: Sung Yool Park. Supervision: Seong Kook Park. Validation: Sung Yool Park. Visualization: Sung Yool Park. Writing—original draft: Sung Yool Park. Writing—review & editing: Seong Kook Park.